Atlas de histología vegetal y animal
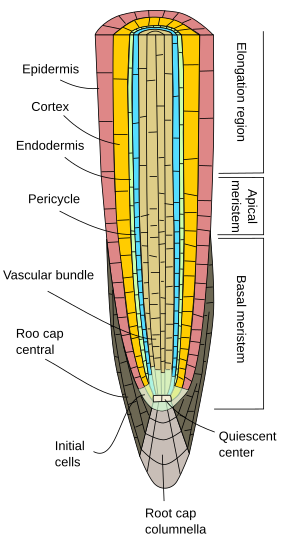
Al principio, las raíces crecen en longitud por la actividad del meristemo apical de la raíz, que está protegido por el capuchón radicular. La proliferación y diferenciación de las células del meristemo apical de la raíz da lugar a células que se organizan primero como la llamada raíz primaria, es decir, presenta un crecimiento primario. Es una organización menos compleja que la del tallo porque carece de nudos, entrenudos y hojas.
Independientemente de la especie vegetal, las raíces primarias muestran una epidermis, o rizodermis, generalmente uniseriada (en capa celular gruesa), carente de estomas, y que muestra pelos radicales en la región de maduración, justo después de la región de elongación (Figuras 1, 2 y 3). En general, cerca de la punta de la raíz, las células epidérmicas muestran una fina cutícula que facilita la entrada de agua y sales minerales. En algunas especies como las plantas xerófitas y en aquellas raíces cercanas a la superficie del suelo, se puede encontrar bajo la epidermis una capa de células conocida como hipodermis. Las células de la hipodermis forman una fina capa de células con paredes celulares suberizadas. La hipodermis puede especializarse aún más y convertirse en la exodermis, que se encuentra en algunas angiospermas, y funciona como una segunda barrera para evitar la difusión libre de sustancias entre la raíz y el suelo.

Los pelos radiculares absorben agua y sales minerales. Son células epidérmicas alargadas que se encuentran en la región de maduración y que aumentan la superficie de absorción en contacto con el medio externo, y por tanto la capacidad de absorción. Los pelos radiculares se forman continuamente y desaparecen a medida que la raíz crece, ya que la región de maduración sigue el crecimiento de la punta de la raíz a una distancia más o menos constante. El número de pelos radiculares es de unos 20 a 500 por cm2 en las raíces de los árboles, y de unos 25000 por cm2 en el centeno de invierno. El número también puede variar en función de las condiciones del entorno.
Hay tres patrones de organización de los pelos radicales (Figura 4). La densidad de los pelos radiculares depende de las condiciones del entorno. Por ejemplo, en suelos con poco fosfato el número de pelos radiculares es mayor para aumentar la superficie total de absorción de las raíces. Muchos organismos simbióticos, como las bacterias fijadoras de nitrógeno, están asociados a los pelos radiculares.


La corteza de la raíz se encuentra debajo de la epidermis o hipodermis. En las raíces, la corteza es gruesa o muy gruesa (mucho más que en los tallos), y generalmente está formada por células de parénquima especializadas en el almacenamiento, aunque puede ser parénquima fotosintético en las raíces aéreas, y parénquima aerífero en las raíces acuáticas. Hay muchos espacios vacíos entre las células de parénquima cortical para que el agua pueda ser conducida (vía apoplástica) hacia el haz vascular. Además, las células de parénquima están conectadas entre sí por plasmodesmos (vía simplástica) que permiten que los minerales y el agua viajen de célula a célula.

Una característica distintiva del crecimiento primario de la raíz es la endodermis, la capa más interna de la corteza. La endodermis se conserva evolutivamente desde los helechos hasta las angiospermas. Es una capa de una célula de espesor de células fuertemente compactadas que tienen paredes celulares parcialmente impregnadas de suberina que forman un engrosamiento, las llamadas rayas de Casparian. La compactación celular y la impermeabilidad de las franjas de Caspary hacen que el agua y las sustancias resueltas atraviesen la endodermis a través del citoplasma de las células de la endodermis. Por lo tanto, la endodermis es una barrera contra la difusión libre que controla las sustancias que viajan desde el suelo hasta los haces vasculares. En aquellas regiones de la raíz con crecimiento secundario, tanto la endodermis como la corteza se pierden (ver página siguiente).
Las rayas casparianas son impregnaciones de la pared primaria que rodean a las células endodérmicas como un cinturón dispuesto longitudinalmente. No son de pared secundaria. Las franjas son continuas entre las células vecinas a través de la laminilla media. En la vista 3D, forman como una red de pesca cilíndrica, donde los cordones son las franjas casparianas y los agujeros son las células de la endodermis. Las franjas casparianas contienen lignina, pero no suberina. Sin embargo, la suberina puede encontrarse en la pared celular de las células de la endodermis después de que se formen las franjas casparianas. La suberina se deposita en forma de capas sobre la superficie de la pared celular, con diferente grosor según la localización de la célula. Por ejemplo, las células de la endodermis que se encuentran más cerca de los polos del xilema no sintetizan suberina. Se denominan células de paso. En algunas especies de plantas, la endodermis experimenta una etapa de desarrollo adicional en la que sus paredes celulares se lignifican y las rayas de Caspio muestran una forma de U. Las células de paso, sin embargo, mantienen paredes celulares delgadas. El continuo crecimiento de las raíces permite la muerte y desaparición de la endodermis.
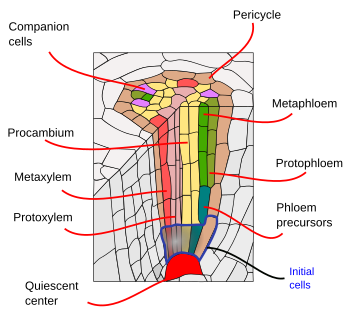
Debajo de la endodermis, hay una o dos capas de células de parénquima con paredes celulares muy finas que forman el periciclo. Las células del periciclo pueden reiniciar la actividad meristemática y formar raíces laterales. En la parte más antigua de las raíces, las células del periciclo están esclerotizadas. En las plantas con raíz secundaria, el periciclo contribuye a la formación del cambium vascular y del felógeno (cambium de corcho).
Los haces vasculares, xilema y floema, se encuentran en la parte interna de la raíz. El xilema primario y el floema primario están dispuestos en filas separadas y alternas. Según el número de filas, hay diarquia (2), triarquia (3), tetrarquia (4). La organización tetrárquica es típica de las dicotiledóneas y las gimnospermas (Figura 5). Las monocotiledóneas son poliarcas que tienen muchas filas de xilema y floema.

Las raíces laterales se generan tras el periodo embrionario y determinan la morfología del sistema radicular de la planta. La formación de la raíz lateral comienza en el periciclo cerca del extremo apical de la raíz, en la región donde las células se diferencian. Esto es diferente de cómo se forman las ramas y las hojas. Las ramas y las hojas se diferencian a partir de los meristemos superficiales, de forma exógena. La localización del punto de inicio de una raíz lateral está influida por la organización del haz vascular. Los primordios de las raíces laterales se forman en puntos opuestos a los polos del xilema en las dicotiledóneas o a los polos del floema en las monocotiledóneas. En algunas especies, la endodermis también colabora en la formación de las raíces laterales.
Las raíces adventicias surgen tras la germinación a partir de células cercanas a los haces vasculares, bien como proceso normal o tras un proceso de inducción. Pueden generarse en tallos, hojas y raíces.
Bibliografía
Furuta KM, Hellmann E, Helariutta Y. 2014. Control molecular de la especificación y diferenciación celular durante el desarrollo procambial. Annual review of plant biology. 65:607-638.
Peret B, De Rybel B, Casimiro I, Benkova E, Swarup R, Laplaze L, Beeckman T, Bennett MJ. 2009. Arabidopsis lateral root development: an emerging story. Trend in plant science. 14: 399-408.
Salazar-Henao J, Vélez-Bermúdez IC, Schmidth W. 2016. La regulación y plasticidad del patrón y la morfogénesis de los pelos de la raíz. Development. 143: 1848-1858.