Biología I

Figura 1. Un nucleótido está formado por tres componentes: una base nitrogenada, un azúcar pentosa y uno o más grupos fosfato. Los residuos de carbono de la pentosa se numeran del 1′ al 5′ (el primo distingue estos residuos de los de la base, que se numeran sin utilizar una notación de primo). La base se une a la posición 1′ de la ribosa, y el fosfato se une a la posición 5′. Cuando se forma un polinucleótido, el fosfato 5′ del nucleótido entrante se une al grupo hidroxilo 3′ al final de la cadena en crecimiento. Hay dos tipos de pentosas en los nucleótidos, la desoxirribosa (presente en el ADN) y la ribosa (presente en el ARN). La desoxirribosa tiene una estructura similar a la de la ribosa, pero tiene un H en lugar de un OH en la posición 2′. Las bases pueden dividirse en dos categorías: purinas y pirimidinas. Las purinas tienen una estructura de doble anillo, y las pirimidinas tienen un solo anillo.
Las bases nitrogenadas, componentes importantes de los nucleótidos, son moléculas orgánicas y se llaman así porque contienen carbono y nitrógeno. Son bases porque contienen un grupo amino que tiene el potencial de unir un hidrógeno extra, y por lo tanto, disminuye la concentración de iones de hidrógeno en su entorno, haciéndolo más básico. Cada nucleótido del ADN contiene una de las cuatro bases nitrogenadas posibles: adenina (A), guanina (G), citosina (C) y timina (T). Los nucleótidos del ARN también contienen una de las cuatro bases posibles: adenina, guanina, citosina y uracilo (U) en lugar de timina.
La adenina y la guanina se clasifican como purinas. La estructura primaria de una purina es de dos anillos de carbono-nitrógeno. La citosina, la timina y el uracilo se clasifican como pirimidinas que tienen un solo anillo de carbono-nitrógeno como estructura primaria (Figura 1). Cada uno de estos anillos básicos de carbono-nitrógeno tiene diferentes grupos funcionales unidos a él. En la taquigrafía de la biología molecular, las bases nitrogenadas se conocen simplemente por sus símbolos A, T, G, C y U. El ADN contiene A, T, G y C, mientras que el ARN contiene A, U, G y C.
El azúcar pentosa en el ADN es la desoxirribosa, y en el ARN, el azúcar es la ribosa (Figura 1). La diferencia entre los azúcares es la presencia del grupo hidroxilo en el segundo carbono de la ribosa y del hidrógeno en el segundo carbono de la desoxirribosa. Los átomos de carbono de la molécula de azúcar se numeran como 1′, 2′, 3′, 4′ y 5′ (1′ se lee como «un primo»). El residuo de fosfato se une al grupo hidroxilo del carbono 5′ de un azúcar y al grupo hidroxilo del carbono 3′ del azúcar del siguiente nucleótido, lo que forma un enlace fosfodiéster 5′-3′. El enlace fosfodiéster no se forma por una simple reacción de deshidratación como los otros enlaces que conectan monómeros en las macromoléculas: su formación implica la eliminación de dos grupos fosfato. Un polinucleótido puede tener miles de estos enlaces fosfodiéster.
Estructura de doble hélice del ADN
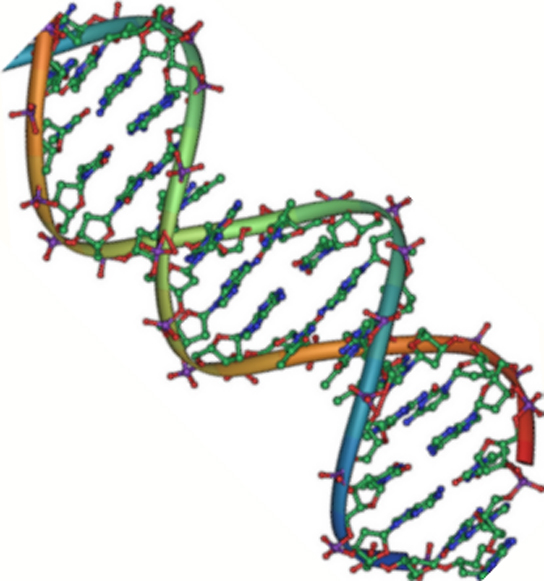
Figura 2. El ADN es una doble hélice antiparalela. La columna vertebral del fosfato (las líneas curvas) está en el exterior, y las bases están en el interior. Cada base interactúa con una base de la cadena opuesta. (crédito: Jerome Walker/Dennis Myts)
Las dos hebras de la hélice van en direcciones opuestas, lo que significa que el extremo del carbono 5′ de una hebra se enfrentará al extremo del carbono 3′ de su hebra correspondiente. (Esto se conoce como orientación antiparalela y es importante para la replicación del ADN y en muchas interacciones de ácidos nucleicos.)
Sólo se permiten ciertos tipos de emparejamiento de bases. Por ejemplo, una determinada purina sólo puede emparejarse con una determinada pirimidina. Esto significa que A puede emparejarse con T, y G puede emparejarse con C, como se muestra en la Figura 3. Esto se conoce como la regla de complementariedad de bases. En otras palabras, las cadenas de ADN son complementarias entre sí. Si la secuencia de una hebra es AATTGGCC, la hebra complementaria tendría la secuencia TTAACCGG. Durante la replicación del ADN, cada hebra se copia, dando lugar a una doble hélice de ADN hija que contiene una hebra de ADN parental y una hebra recién sintetizada.
Práctica
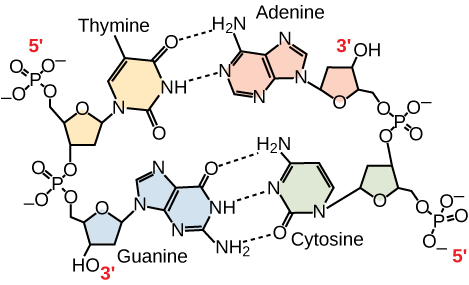
Figura 3. En una molécula de ADN de doble hebra, las dos hebras discurren antiparalelas entre sí, de modo que una hebra va de 5′ a 3′ y la otra de 3′ a 5′. La espina dorsal de fosfato se encuentra en el exterior y las bases en el centro. La adenina forma enlaces de hidrógeno (o pares de bases) con la timina, y la guanina pares de bases con la citosina.
Se produce una mutación, y la citosina se sustituye por adenina. ¿Qué impacto crees que tendrá esto en la estructura del ADN?
ARN
El ácido ribonucleico, o ARN, participa principalmente en el proceso de síntesis de proteínas bajo la dirección del ADN. El ARN suele ser monocatenario y está formado por ribonucleótidos unidos por enlaces fosfodiéster. Un ribonucleótido de la cadena de ARN contiene ribosa (el azúcar pentosa), una de las cuatro bases nitrogenadas (A, U, G y C) y el grupo fosfato.
Hay cuatro tipos principales de ARN: el ARN mensajero (ARNm), el ARN ribosomal (ARNr), el ARN de transferencia (ARNt) y el microARN (ARNm). El primero, el ARNm, transporta el mensaje del ADN, que controla todas las actividades celulares de una célula. Si una célula necesita que se sintetice una determinada proteína, el gen de este producto se «enciende» y el ARN mensajero se sintetiza en el núcleo. La secuencia de bases del ARN es complementaria a la secuencia codificadora del ADN del que se ha copiado. Sin embargo, en el ARN, la base T está ausente y en su lugar está presente la U. Si la cadena de ADN tiene una secuencia AATTGCGC, la secuencia del ARN complementario es UUAACGCG. En el citoplasma, el ARNm interactúa con los ribosomas y otra maquinaria celular (Figura 4).
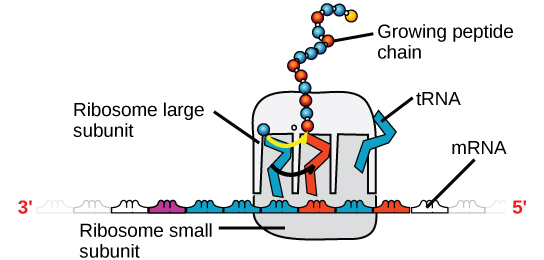
Figura 4. Un ribosoma tiene dos partes: una subunidad grande y una subunidad pequeña. El ARNm se sitúa entre las dos subunidades. Una molécula de ARNt reconoce un codón en el ARNm, se une a él mediante el emparejamiento de bases complementarias y añade el aminoácido correcto a la cadena peptídica en crecimiento.
El ARNm se lee en conjuntos de tres bases conocidos como codones. Cada codón codifica un único aminoácido. De este modo, se lee el ARNm y se fabrica el producto proteico. El ARN ribosómico (ARNr) es uno de los principales componentes de los ribosomas en los que se une el ARNm. El ARNr asegura la correcta alineación del ARNm y los ribosomas; el ARNr del ribosoma también tiene una actividad enzimática (peptidil transferasa) y cataliza la formación de los enlaces peptídicos entre dos aminoácidos alineados. El ARN de transferencia (ARNt) es uno de los más pequeños de los cuatro tipos de ARN, normalmente de 70-90 nucleótidos. Lleva el aminoácido correcto al lugar de la síntesis de proteínas. Es el emparejamiento de bases entre el ARNt y el ARNm lo que permite que el aminoácido correcto se inserte en la cadena polipeptídica. Los microARN son las moléculas de ARN más pequeñas y su función consiste en la regulación de la expresión de los genes al interferir en la expresión de determinados mensajes de ARNm.