Expresiones moleculares Biología celular: Filamentos intermedios
Filamentos intermedios
Los filamentos intermedios son una clase muy amplia de proteínas fibrosas que desempeñan un papel importante como elementos estructurales y funcionales del citoesqueleto. Con un tamaño que oscila entre los 8 y los 12 nanómetros (de diámetro; véase la Figura 1), los filamentos intermedios funcionan como elementos portadores de tensión para ayudar a mantener la forma y la rigidez de la célula, y sirven para anclar en su lugar varios orgánulos, incluidos el núcleo y los desmosomas. Los filamentos intermedios también participan en la formación de la lámina nuclear, un conjunto de malla similar a una red que recubre la membrana nuclear interna y gobierna la forma del núcleo.
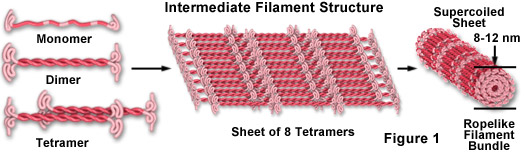
Aunque todos los eucariotas contienen los elementos comunes del citoesqueleto actina y tubulina (ambos libres en el citoplasma y polimerizados en forma de microfilamentos y microtúbulos), Los filamentos intermedios sólo se encuentran en algunas especies de metazoos, como los vertebrados, los nematodos y los moluscos. En los vertebrados, la presencia y composición de los filamentos intermedios no sólo depende de la especie, sino que también varía según el tipo de tejido. Por ejemplo, la mayoría de las células epiteliales de los animales contienen queratinas, una familia diversa de filamentos intermedios que consta de más de 50 miembros, mientras que las células mesenquimales y musculares son ricas en las proteínas fibrosas vimentina y desmina, respectivamente. Los filamentos intermedios que se encuentran en las neuronas y las células gliales incluyen la periferina, los neurofilamentos y la proteína ácida fibrilar glial (GFAP). Una variedad de proteínas asociadas se unen a los filamentos intermedios, ya sea para mejorar la estabilidad (a través de la reticulación) o para proporcionar sitios de unión para otros conjuntos de proteínas, tales como los filamentos de actina y los microtúbulos.
Otra clase altamente especializada de filamentos intermedios son las láminas nucleares, que constituyen la red de proteínas fibrosas que recubren el interior de la membrana nuclear, como se discutió anteriormente. Debido a su proximidad a la membrana, las láminas nucleares ayudan a unir los cromosomas a la membrana nuclear y proporcionan puntos de anclaje para los poros nucleares. Muchos científicos creen que las láminas nucleares son el ancestro evolutivo de los filamentos intermedios citoplasmáticos, que evolucionaron mediante la duplicación y la translocación del producto génico al citoplasma. La rigidez que otorgan a las células las redes de filamentos intermedios es especialmente útil para los animales de cuerpo blando que no poseen un exoesqueleto. Dado que los filamentos intermedios son muy abundantes en las células que suelen estar sometidas a una gran tensión mecánica in vivo, parece que su función principal es proporcionar fuerza física a las células y los tejidos.
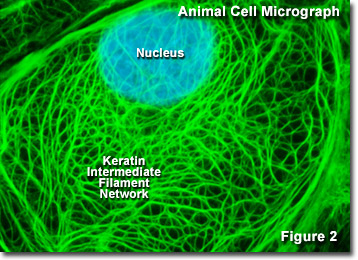
En la Figura 2 se presenta una imagen digital de la red intermedia de queratina encontrada en una célula epitelial de canguro de rata (línea PtK2) vista a través de un microscopio óptico de fluorescencia. La extensa red entrelazada se marcó con anticuerpos primarios contra varias proteínas de citoqueratina, que luego se tiñeron con anticuerpos secundarios que contenían un colorante verde fluorescente. El núcleo se contrateñó con un colorante azul para observar su ubicación en relación con la red de filamentos intermedios. La microscopía de fluorescencia es una herramienta importante que los científicos utilizan para examinar la estructura y la función de los orgánulos celulares internos y el citoesqueleto.
Como se ilustra en la Figura 1, los péptidos monoméricos de filamentos intermedios son una clase de proteínas fibrosas alargadas con una región central alfa-hélica coronada con extremos globulares tanto en el extremo amino como en el carboxílico. Dos de las unidades monoméricas forman un dímero en espiral que se autoasocian en una disposición antiparalela para formar un tetrámero escalonado, que es la subunidad soluble análoga para el monómero globular de actina y el heterodímero de tubulina (que existe libre en el citoplasma). Las unidades del tetrámero se empaquetan lateralmente para formar una hoja de ocho protofilamentos paralelos que están superenrollados en un haz apretado. La sección transversal de cada filamento intermedio fuertemente enrollado revela 32 péptidos alfa-helicoidales individuales, lo que hace que el filamento sea fácil de doblar pero bastante difícil de romper, lo que explica su extrema rigidez estructural. Aunque se sabe menos sobre el mecanismo de ensamblaje y desensamblaje de los filamentos intermedios, está claro que algunas clases son estructuras muy dinámicas con una tasa significativa de recambio en muchos tipos de células.
Las mutaciones en los genes de los filamentos intermedios dan lugar a una serie de enfermedades poco comunes. Por ejemplo, las queratinas defectuosas en el tejido cutáneo dan lugar a un trastorno conocido como epidermólisis bullosa simple, que se manifiesta con ampollas en la piel producidas incluso con un ligero estrés mecánico. Otras enfermedades ampollosas similares debidas a mutaciones de las queratinas en otros tejidos afectan al esófago, los ojos y la boca. Varias enfermedades neurodegenerativas, como la esclerosis lateral amiotrófica (ELA o enfermedad de Lou Gehrig), se asocian a fallos en la red de filamentos intermedios (neurofilamentos), y los defectos en los filamentos intermedios de desmina producen trastornos musculares.
Volver a la estructura celular animal
Volver a la estructura celular vegetal
¿Preguntas o comentarios? Envíanos un correo electrónico.
© 1995-2021 por Michael W. Davidson y The Florida State University. Todos los derechos reservados. No se pueden reproducir ni utilizar imágenes, gráficos, software, scripts o applets de ninguna manera sin el permiso de los titulares de los derechos de autor. El uso de este sitio web significa que usted está de acuerdo con todos los Términos y Condiciones Legales establecidos por los propietarios.
Este sitio web es mantenido por nuestro
Graphics & Equipo de Programación Web
en colaboración con Microscopía Óptica en el
Laboratorio Nacional de Alto Campo Magnético.
Última modificación: Friday, Nov 13, 2015 at 02:18 PM
Cuento de accesos desde el 14 de diciembre de 2004: 173927
Microscopios proporcionados por:
![]()
![]()