Atlas d’histologie végétale et animale
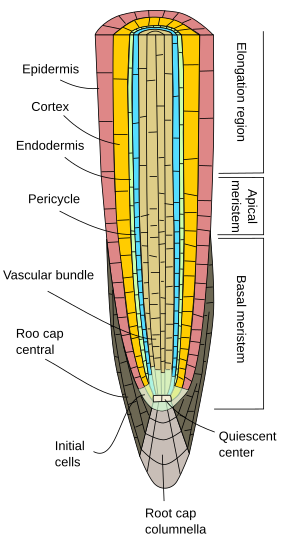
Initialement, les racines croissent en longueur par l’activité du méristème apical racinaire, qui est protégé par le chapeau racinaire. La prolifération et la différenciation des cellules du méristème apical des racines donnent des cellules qui s’organisent d’abord comme la racine dite primaire, c’est-à-dire qu’elle présente une croissance primaire. C’est une organisation moins complexe que la tige car elle est dépourvue de nœuds, d’entre-nœuds et de feuilles.
Quelle que soit l’espèce végétale, les racines primaires présentent un épiderme, ou rhyzoderme, généralement uniseri (sur une couche épaisse de cellules), dépourvu de stomates, et présentant des poils racinaires dans la région de maturation, juste après la région d’élongation (Figures 1, 2 et 3). En général, près de l’extrémité de la racine, les cellules épidermiques présentent une cuticule fine facilitant l’entrée de l’eau et des sels minéraux. Chez certaines espèces comme les plantes xérophytes et dans les racines proches de la surface du sol, on trouve sous l’épiderme une couche de cellules appelée hypoderme. Les cellules de l’hypoderme forment une fine couche de cellules aux parois cellulaires subérisées. L’hypoderme peut se spécialiser davantage et devenir l’exoderme, que l’on trouve chez certains angiospermes, et qui fonctionne comme une deuxième barrière pour empêcher la libre diffusion des substances entre la racine et le sol.

Les poils racinaires absorbent l’eau et les sels minéraux. Ce sont des cellules épidermiques allongées présentes dans la région de maturation qui augmentent la surface d’absorption en contact avec le milieu extérieur, et donc la capacité d’absorption. Les poils racinaires se forment et disparaissent continuellement au cours de la croissance de la racine car la région de maturation suit la croissance de l’extrémité de la racine à une distance plus ou moins constante. Le nombre de poils racinaires est d’environ 20 à 500 par cm2 dans les racines des arbres, et d’environ 25000 par cm2 dans le seigle d’hiver. Ce nombre peut également varier en fonction des conditions de l’environnement.
Il existe trois modes d’organisation des poils racinaires (figure 4). La densité des poils racinaires dépend des conditions de l’environnement. Par exemple, dans les sols pauvres en phosphates, le nombre de poils racinaires est plus élevé pour augmenter la surface totale d’absorption des racines. De nombreux organismes symbiotiques, tels que les bactéries fixatrices d’azote, sont associés aux poils racinaires.


Le cortex des racines se trouve sous l’épiderme, ou hypoderme. Dans les racines, le cortex est épais ou très épais (beaucoup plus que dans les tiges), et il est généralement constitué de cellules de parenchyme spécialisées dans le stockage, bien qu’il puisse s’agir de parenchyme photosynthétique dans les racines aériennes, et de parenchyme aéré dans les racines aquatiques. Il existe de nombreux espaces vides entre les cellules du parenchyme cortical afin que l’eau puisse être conduite (voie apoplastique) vers le faisceau vasculaire. De plus, les cellules du parenchyme sont reliées entre elles par des plasmodesmes (voie symplastique) qui permettent aux minéraux et à l’eau de voyager de cellule en cellule.

Une caractéristique distincte de la croissance primaire des racines est l’endoderme, la couche la plus interne du cortex. L’endoderme est conservé au cours de l’évolution, des fougères aux angiospermes. Il s’agit d’une couche d’une cellule d’épaisseur composée de cellules étroitement compactes dont les parois cellulaires sont partiellement imprégnées de subérine et qui forment un épaississement, les fameuses bandes caspiennes. La compaction cellulaire et l’imperméabilité des bandes caspariens font que l’eau et les substances dissoutes traversent l’endoderme par le cytoplasme des cellules de l’endoderme. Ainsi, l’endoderme est une barrière contre la diffusion libre qui contrôle les substances voyageant du sol vers les faisceaux vasculaires. Dans les régions racinaires présentant une croissance secondaire, l’endoderme et le cortex sont tous deux perdus (voir page suivante).
Les bandes caspiennes sont des imprégnations de la paroi primaire qui entourent les cellules endodermiques sous forme d’une ceinture disposée longitudinalement. Elles ne constituent pas une paroi secondaire. Les bandes sont continues entre les cellules voisines à travers la lamelle moyenne. En vue 3D, elles forment comme un filet de pêche cylindrique, où les cordons sont les bandes caspariens et les trous sont les cellules endodermiques. Les bandes caspariens contiennent de la lignine, mais pas de subérine. Cependant, la subérine peut être trouvée dans la paroi cellulaire des cellules de l’endoderme après la formation des bandes caspariales. La subérine est déposée en couches sur la surface de la paroi cellulaire, avec une épaisseur différente selon la localisation de la cellule. Par exemple, les cellules de l’endoderme situées plus près des pôles du xylème ne synthétisent pas de subérine. Elles sont appelées cellules de passage. Chez certaines espèces végétales, l’endoderme subit un stade de développement supplémentaire au cours duquel ses parois cellulaires se lignifient et les bandes caspiennes présentent une forme en U. Les cellules de passage, cependant, conservent des parois cellulaires fines. La croissance continue des racines laisse à la mort et à la disparition de l’endoderme.
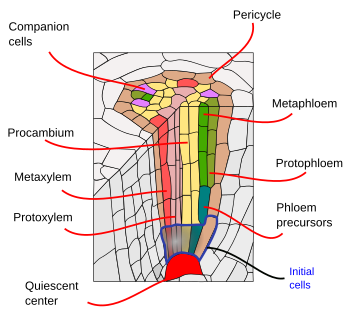
Sous l’endoderme, il y a une ou deux couches de cellules de parenchyme avec des parois cellulaires très fines qui forment le péricycle. Les cellules du péricycle peuvent relancer l’activité méristématique et former des racines latérales. Dans la partie la plus ancienne des racines, les cellules du péricycle sont sclérosées. Chez les plantes à racine secondaire, le péricycle contribue à la formation du cambium vasculaire et du phellogène (cambium de liège).
Les faisceaux vasculaires, xylème et phloème, se trouvent dans la partie interne de la racine. Le xylème primaire et le phloème primaire sont disposés en rangées séparées et alternées. Selon le nombre de rangées, on distingue les diarches (2), triarches (3), tétrarches (4). L’organisation en tétrarch est typique des dicotylédones et des gimnospermes (Figure 5). Les monocotylédones sont polyarches ayant de nombreuses rangées de xylème et de phloème.

Les racines latérales sont générées après la période embryonnaire et déterminent la morphologie du système racinaire de la plante. La formation des racines latérales commence dans le péricycle près de l’extrémité apicale de la racine, dans la région où les cellules se différencient. Ce processus est différent de celui de la formation des branches et des feuilles. Les branches et les feuilles se différencient à partir de méristèmes superficiels, de manière exogène. L’emplacement du point de départ d’une racine latérale est influencé par l’organisation du faisceau vasculaire. Les primordia des racines latérales se forment en des points opposés aux pôles du xylème chez les dicotylédones ou aux pôles du phloème chez les monocotylédones. Chez certaines espèces, l’endoderme collabore également à la formation des racines latérales.
Les racines adventives émergent après la germination à partir de cellules proches des faisceaux vasculaires, soit comme un processus normal, soit après un processus d’induction. Elles peuvent être générées dans les tiges, les feuilles et les racines.
Bibliographie
Furuta KM, Hellmann E, Helariutta Y. 2014. Contrôle moléculaire de la spécification et de la différenciation cellulaire au cours du développement procambial. Revue annuelle de biologie végétale. 65:607-638.
Peret B, De Rybel B, Casimiro I, Benkova E, Swarup R, Laplaze L, Beeckman T, Bennett MJ. 2009. Le développement des racines latérales d’Arabidopsis : une histoire émergente. Tendance dans la science des plantes. 14 : 399-408.
Salazar-Henao J, Vélez-Bermúdez IC, Schmidth W. 2016. La régulation et la plasticité du patterning et de la morphogenèse du chevelu racinaire. Développement. 143 : 1848-1858.