Atlante di istologia vegetale e animale
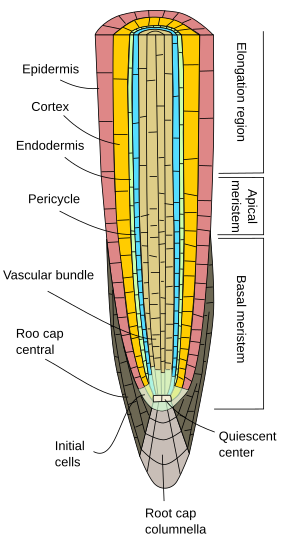
Inizialmente, le radici crescono in lunghezza grazie all’attività del meristema apicale radicale, che è protetto dal cappello radicale. La proliferazione e la differenziazione delle cellule del meristema apicale della radice danno cellule che si organizzano dapprima come la cosiddetta radice primaria, cioè, mostra una crescita primaria. Si tratta di un’organizzazione meno complessa del fusto perché manca di nodi, internodi e foglie.
Non importa la specie di pianta, le radici primarie mostrano un’epidermide, o rizoderma, di solito uniseriata (su strato spesso di cellule), priva di stomi, e che mostra peli radicali nella regione di maturazione, subito dopo la regione di allungamento (figure 1, 2 e 3). In generale, vicino alla punta della radice, le cellule epidermiche mostrano una sottile cuticola che facilita l’ingresso di acqua e sali minerali. In alcune specie come le piante xerofite e in quelle radici vicine alla superficie del suolo, uno strato di cellule noto come ipoderma può essere trovato sotto l’epidermide. Le cellule dell’ipoderma formano un sottile strato di cellule con pareti cellulari suberizzate. L’ipoderma può essere ulteriormente specializzato e diventare l’esoderma, che si trova in alcune angiosperme, e funziona come una seconda barriera per impedire la libera diffusione delle sostanze tra la radice e il suolo.

I peli radicali assorbono acqua e sali minerali. Sono cellule epidermiche allungate che si trovano nella regione di maturazione e che aumentano la superficie di assorbimento a contatto con l’ambiente esterno, e quindi la capacità di assorbimento. I peli radicali si formano e scompaiono continuamente durante la crescita della radice perché la regione di maturazione segue la crescita della punta della radice ad una distanza più o meno costante. Il numero di peli radicali è di circa 20-500 per cm2 nelle radici degli alberi, e di circa 25000 per cm2 nella segale invernale. Il numero può anche variare a seconda delle condizioni ambientali.
Ci sono tre modelli di organizzazione dei peli radicali (Figura 4). La densità dei peli radicali dipende dalle condizioni ambientali. Per esempio, in terreni a basso contenuto di fosfati, il numero di peli radicali è maggiore per aumentare la superficie totale di assorbimento delle radici. Molti organismi simbiotici, come i batteri che fissano l’azoto, sono associati ai peli radicali.


La corteccia della radice si trova sotto l’epidermide, o ipoderma. Nelle radici, la corteccia è spessa o molto spessa (molto più che nei fusti), ed è generalmente fatta di cellule parenchimali specializzate nell’immagazzinamento, anche se può essere parenchima fotosintetico nelle radici aeree, e parenchima aerifero nelle radici acquatiche. Ci sono molti spazi vuoti tra le cellule del parenchima corticale in modo che l’acqua possa essere condotta (via apoplastica) verso il fascio vascolare. Inoltre, le cellule del parenchima sono collegate tra loro da plasmodesmata (via simplastica) che permettono ai minerali e all’acqua di viaggiare da cellula a cellula.

Una caratteristica distinta della crescita primaria delle radici è l’endoderma, lo strato più interno della corteccia. L’endoderma è evolutivamente conservato dalle felci alle angiosperme. È uno strato spesso una cella di cellule strettamente compattate con pareti cellulari parzialmente impregnate di suberina che formano un ispessimento, le cosiddette strisce di Casparian. La compattazione cellulare e l’impermeabilità delle strisce di Caspary fanno sì che l’acqua e le sostanze risolte attraversino l’endoderma attraverso il citoplasma delle cellule dell’endoderma. Quindi, l’endoderma è una barriera contro la libera diffusione che controlla le sostanze che viaggiano dal suolo ai fasci vascolari. In quelle regioni di radici con crescita secondaria, sia l’endoderma che la corteccia sono persi (vedi pagina seguente).
Le strisce di Caspari sono impregnazioni della parete primaria che circondano le cellule endodermiche come una cintura disposta longitudinalmente. Non sono pareti secondarie. Le strisce sono continue tra cellule vicine attraverso la lamella centrale. In vista 3D, formano come una rete da pesca cilindrica, dove i cordoni sono le strisce di Caspari e i fori sono le cellule dell’endoderma. Le strisce di Casparian contengono lignina, ma non suberina. Tuttavia, la suberina può essere trovata nella parete cellulare delle cellule endodermiche dopo che le strisce di Caspari sono formate. La suberina è depositata come strati sulla superficie della parete cellulare, con spessore diverso a seconda della localizzazione della cellula. Per esempio, le cellule dell’endoderma che si trovano più vicine ai poli dello xilema non sintetizzano la suberina. Sono chiamate cellule di passaggio. In alcune specie di piante, l’endoderma subisce un ulteriore stadio di sviluppo in cui le loro pareti cellulari diventano lignificate e le strisce di Casparian mostrano una forma a U. Le cellule di passaggio, tuttavia, mantengono pareti cellulari sottili. La continua crescita delle radici porta alla morte e alla scomparsa dell’endoderma.
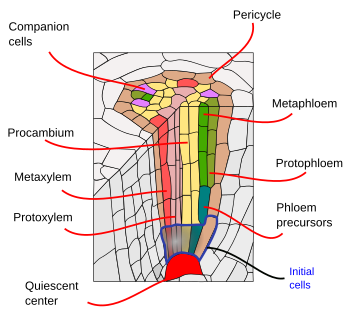
Sotto l’endoderma, ci sono uno o due strati di cellule del parenchima con pareti cellulari molto sottili che formano il periciclo. Le cellule del periciclo possono riprendere l’attività meristematica e formare radici laterali. Nella parte più vecchia delle radici, le cellule del periciclo sono sclerotizzate. Nelle piante con radice secondaria, il periciclo contribuisce alla formazione del cambio vascolare e del fellogeno (cambio di sughero).
I fasci vascolari, xilema e floema, si trovano nella parte interna della radice. Lo xilema primario e il floema primario sono disposti in file separate e alternate. Secondo il numero di file, ci sono diarca (2), triarca (3), tetrarca (4). L’organizzazione tetrarca è tipica delle dicotiledoni e delle Gimnosperme (Figura 5). Le monocotiledoni sono poliarchiche con molte file di xilema e floema.

Le radici laterali sono generate dopo il periodo embrionale e determinano la morfologia del sistema radicale della pianta. La formazione delle radici laterali inizia nel periciclo vicino alla punta apicale della radice, nella regione in cui le cellule si differenziano. Questo è diverso da come si formano i rami e le foglie. I rami e le foglie si differenziano dai meristemi superficiali, in modo esogeno. La posizione del punto di partenza di una radice laterale è influenzata dall’organizzazione del fascio vascolare. I primordi delle radici laterali si formano in punti opposti ai poli dello xilema nelle dicotiledoni o ai poli del floema nelle monocotiledoni. In alcune specie, anche l’endoderma collabora alla formazione delle radici laterali.
Le radici avventizie emergono dopo la germinazione da cellule vicine ai fasci vascolari, come un processo normale o dopo un processo di induzione. Possono essere generate in steli, foglie e radici.
Bibliografia
Furuta KM, Hellmann E, Helariutta Y. 2014. Controllo molecolare della specificazione e differenziazione cellulare durante lo sviluppo procambico. Rassegna annuale di biologia vegetale. 65:607-638.
Peret B, De Rybel B, Casimiro I, Benkova E, Swarup R, Laplaze L, Beeckman T, Bennett MJ. 2009. Arabidopsis sviluppo della radice laterale: una storia emergente. Tendenza nella scienza delle piante. 14: 399-408.
Salazar-Henao J, Vélez-Bermúdez IC, Schmidth W. 2016. La regolazione e la plasticità del patterning e della morfogenesi dei peli delle radici. Sviluppo. 143: 1848-1858.