Potenziale di membrana
Il potenziale di membrana in una cellula deriva in definitiva da due fattori: forza elettrica e diffusione. La forza elettrica deriva dall’attrazione reciproca tra particelle con cariche elettriche opposte (positive e negative) e la repulsione reciproca tra particelle con lo stesso tipo di carica (entrambe positive o entrambe negative). La diffusione deriva dalla tendenza statistica delle particelle a ridistribuirsi da regioni dove sono altamente concentrate a regioni dove la concentrazione è bassa.
VoltageEdit
La tensione, che è sinonimo di differenza di potenziale elettrico, è la capacità di guidare una corrente elettrica attraverso una resistenza. Infatti, la definizione più semplice di una tensione è data dalla legge di Ohm: V=IR, dove V è la tensione, I è la corrente e R è la resistenza. Se una fonte di tensione come una batteria viene inserita in un circuito elettrico, maggiore è la tensione della fonte, maggiore è la quantità di corrente che guiderà attraverso la resistenza disponibile. Il significato funzionale della tensione risiede solo nelle differenze di potenziale tra due punti in un circuito. L’idea di una tensione in un singolo punto non ha senso. È convenzionale in elettronica assegnare una tensione di zero a qualche elemento del circuito scelto arbitrariamente, e poi assegnare tensioni per altri elementi misurati rispetto a quel punto zero. Non c’è alcun significato in quale elemento viene scelto come punto zero – la funzione di un circuito dipende solo dalle differenze, non dalle tensioni in sé. Tuttavia, nella maggior parte dei casi e per convenzione, il livello zero è più spesso assegnato alla porzione di un circuito che è in contatto con la terra.
Lo stesso principio si applica alla tensione nella biologia cellulare. Nei tessuti elettricamente attivi, la differenza di potenziale tra due punti qualsiasi può essere misurata inserendo un elettrodo in ogni punto, per esempio uno all’interno e uno all’esterno della cellula, e collegando entrambi gli elettrodi ai cavi di quello che è in sostanza un voltmetro specializzato. Per convenzione, il valore zero del potenziale è assegnato all’esterno della cella e il segno della differenza di potenziale tra l’esterno e l’interno è determinato dal potenziale dell’interno rispetto allo zero esterno.
In termini matematici, la definizione di tensione inizia con il concetto di campo elettrico E, un campo vettoriale che assegna una grandezza e una direzione a ogni punto nello spazio. In molte situazioni, il campo elettrico è un campo conservativo, il che significa che può essere espresso come il gradiente di una funzione scalare V, cioè E = -∇V. Questo campo scalare V è indicato come la distribuzione della tensione. Si noti che la definizione permette una costante d’integrazione arbitraria – ecco perché i valori assoluti di tensione non sono significativi. In generale, i campi elettrici possono essere trattati come conservativi solo se i campi magnetici non li influenzano significativamente, ma questa condizione di solito si applica bene ai tessuti biologici.
Perché il campo elettrico è il gradiente della distribuzione di tensione, rapidi cambiamenti di tensione all’interno di una piccola regione implicano un forte campo elettrico; al contrario, se la tensione rimane approssimativamente la stessa in una grande regione, i campi elettrici in quella regione devono essere deboli. Un forte campo elettrico, equivalente a un forte gradiente di tensione, implica che una forte forza viene esercitata su qualsiasi particella carica che si trova all’interno della regione.
Gli ioni e le forze che guidano il loro movimentoModifica

I segnali elettrici all’interno degli organismi biologici sono, in generale, guidati dagli ioni. I cationi più importanti per il potenziale d’azione sono il sodio (Na+) e il potassio (K+). Entrambi sono cationi monovalenti che portano una singola carica positiva. I potenziali d’azione possono anche coinvolgere il calcio (Ca2+), che è un catione divalente che porta una doppia carica positiva. L’anione cloruro (Cl-) gioca un ruolo importante nei potenziali d’azione di alcune alghe, ma gioca un ruolo trascurabile nei potenziali d’azione della maggior parte degli animali.
Gli ioni attraversano la membrana cellulare sotto due influenze: diffusione e campi elettrici. Un semplice esempio in cui due soluzioni -A e B- sono separate da una barriera porosa illustra che la diffusione farà sì che alla fine si mescolino in soluzioni uguali. Questo mescolamento avviene a causa della differenza delle loro concentrazioni. La regione ad alta concentrazione si diffonderà verso la regione a bassa concentrazione. Per estendere l’esempio, lasciamo che la soluzione A abbia 30 ioni di sodio e 30 ioni di cloruro. Inoltre, lasciamo che la soluzione B abbia solo 20 ioni sodio e 20 ioni cloruro. Supponendo che la barriera permetta a entrambi i tipi di ioni di attraversarla, allora si raggiungerà uno stato stazionario in cui entrambe le soluzioni hanno 25 ioni sodio e 25 ioni cloruro. Se, tuttavia, la barriera porosa è selettiva a quali ioni sono lasciati passare, allora la diffusione da sola non determinerà la soluzione risultante. Tornando all’esempio precedente, costruiamo ora una barriera che sia permeabile solo agli ioni di sodio. Ora, solo al sodio è permesso di diffondere attraverso la barriera dalla sua più alta concentrazione nella soluzione A alla più bassa concentrazione nella soluzione B. Questo risulterà in un maggiore accumulo di ioni sodio rispetto agli ioni cloruro nella soluzione B e un minor numero di ioni sodio rispetto agli ioni cloruro nella soluzione A.
Questo significa che c’è una carica positiva netta nella soluzione B dalla maggiore concentrazione di ioni sodio caricati positivamente rispetto agli ioni cloruro caricati negativamente. Allo stesso modo, c’è una carica netta negativa nella soluzione A dalla maggiore concentrazione di ioni cloruro negativi rispetto agli ioni di sodio positivi. Poiché le cariche opposte si attraggono e le cariche simili si respingono, gli ioni sono ora anche influenzati dai campi elettrici e dalle forze di diffusione. Pertanto, gli ioni di sodio positivi avranno meno probabilità di viaggiare verso la soluzione B, ora più positiva, e di rimanere nella soluzione A, ora più negativa. Il punto in cui le forze dei campi elettrici contrastano completamente la forza dovuta alla diffusione è chiamato potenziale di equilibrio. A questo punto, il flusso netto dello ione specifico (in questo caso il sodio) è zero.
Membrane plasmaticheModifica

Ogni cellula è racchiusa in una membrana plasmatica, che ha la struttura di un bilayer lipidico con molti tipi di grandi molecole incorporate in essa. Essendo fatta di molecole lipidiche, la membrana plasmatica ha intrinsecamente un’alta resistività elettrica, in altre parole una bassa permeabilità intrinseca agli ioni. Tuttavia, alcune delle molecole incorporate nella membrana sono in grado di trasportare attivamente gli ioni da un lato all’altro della membrana o di fornire canali attraverso i quali essi possono muoversi.
In terminologia elettrica, la membrana plasmatica funziona come una resistenza e un condensatore combinati. La resistenza deriva dal fatto che la membrana impedisce il movimento delle cariche attraverso di essa. La capacità deriva dal fatto che il bilayer lipidico è così sottile che un accumulo di particelle cariche su un lato dà origine a una forza elettrica che tira le particelle con carica opposta verso l’altro lato. La capacità della membrana non è relativamente influenzata dalle molecole che vi sono incorporate, quindi ha un valore più o meno invariante stimato a circa 2 μF/cm2 (la capacità totale di un pezzo di membrana è proporzionale alla sua area). La conduttanza di un doppio strato lipidico puro è così bassa, d’altra parte, che in situazioni biologiche è sempre dominata dalla conduttanza delle vie alternative fornite dalle molecole incorporate. Così, la capacità della membrana è più o meno fissa, ma la resistenza è molto variabile.
Lo spessore di una membrana plasmatica è stimato in circa 7-8 nanometri. Poiché la membrana è così sottile, non ci vuole una tensione transmembrana molto grande per creare un forte campo elettrico al suo interno. I tipici potenziali di membrana nelle cellule animali sono dell’ordine di 100 millivolt (cioè un decimo di volt), ma i calcoli mostrano che questo genera un campo elettrico vicino al massimo che la membrana può sostenere – è stato calcolato che una differenza di tensione molto più grande di 200 millivolt potrebbe causare una rottura dielettrica, cioè un arco attraverso la membrana.
Diffusione e trasporto facilitatoModifica

La resistenza di un bilayer lipidico puro al passaggio di ioni attraverso di esso è molto alta, ma le strutture incorporate nella membrana possono migliorare notevolmente il movimento degli ioni, sia attivamente che passivamente, attraverso meccanismi chiamati trasporto facilitato e diffusione facilitata. I due tipi di strutture che giocano i ruoli più importanti sono i canali ionici e le pompe ioniche, entrambi solitamente formati da assemblaggi di molecole proteiche. I canali ionici forniscono passaggi attraverso i quali gli ioni possono muoversi. Nella maggior parte dei casi, un canale ionico è permeabile solo a specifici tipi di ioni (per esempio, sodio e potassio ma non cloruro o calcio), e a volte la permeabilità varia a seconda della direzione del movimento ionico. Le pompe ioniche, conosciute anche come trasportatori di ioni o proteine trasportatrici, trasportano attivamente tipi specifici di ioni da un lato all’altro della membrana, a volte usando energia derivata da processi metabolici per farlo.
Pompe ionicheModifica
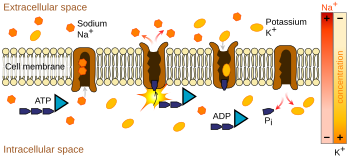
Le pompe ioniche sono proteine integrali di membrana che effettuano il trasporto attivo, cioè usano l’energia cellulare (ATP) per “pompare” gli ioni contro il loro gradiente di concentrazione. Tali pompe ioniche prendono ioni da un lato della membrana (diminuendone la concentrazione) e li rilasciano dall’altro lato (aumentandone la concentrazione).
La pompa ionica più rilevante per il potenziale d’azione è la pompa sodio-potassio, che trasporta tre ioni sodio fuori dalla cellula e due ioni potassio dentro. Di conseguenza, la concentrazione di ioni potassio K+ all’interno del neurone è circa 20 volte più grande della concentrazione esterna, mentre la concentrazione di sodio all’esterno è circa nove volte più grande dell’interno. In modo simile, altri ioni hanno concentrazioni diverse all’interno e all’esterno del neurone, come calcio, cloruro e magnesio.
Se i numeri di ogni tipo di ione fossero uguali, la pompa sodio-potassio sarebbe elettricamente neutra, ma, a causa dello scambio tre per due, dà un movimento netto di una carica positiva da intracellulare a extracellulare per ogni ciclo, contribuendo così ad una differenza di tensione positiva. La pompa ha tre effetti: (1) rende la concentrazione di sodio alta nello spazio extracellulare e bassa nello spazio intracellulare; (2) rende la concentrazione di potassio alta nello spazio intracellulare e bassa nello spazio extracellulare; (3) dà allo spazio intracellulare una tensione negativa rispetto allo spazio extracellulare.
La pompa sodio-potassio è relativamente lenta nel funzionamento. Se una cellula fosse inizializzata con concentrazioni uguali di sodio e potassio ovunque, ci vorrebbero ore perché la pompa stabilisca l’equilibrio. La pompa funziona costantemente, ma diventa progressivamente meno efficiente man mano che si riducono le concentrazioni di sodio e potassio disponibili per il pompaggio.
Le pompe ioniche influenzano il potenziale d’azione solo stabilendo il rapporto relativo delle concentrazioni di ioni intracellulari ed extracellulari. Il potenziale d’azione coinvolge principalmente l’apertura e la chiusura dei canali ionici, non le pompe ioniche. Se le pompe ioniche vengono spente rimuovendo la loro fonte di energia o aggiungendo un inibitore come l’ouabain, l’assone può ancora sparare centinaia di migliaia di potenziali d’azione prima che la loro ampiezza cominci a decadere significativamente. In particolare, le pompe ioniche non giocano un ruolo significativo nella ripolarizzazione della membrana dopo un potenziale d’azione.
Un’altra pompa ionica funzionalmente importante è lo scambiatore sodio-calcio. Questa pompa funziona in modo concettualmente simile alla pompa sodio-potassio, tranne che in ogni ciclo scambia tre Na+ dallo spazio extracellulare per un Ca++ dallo spazio intracellulare. Poiché il flusso netto di carica è verso l’interno, questa pompa funziona “in discesa”, in effetti, e quindi non richiede alcuna fonte di energia tranne la tensione di membrana. Il suo effetto più importante è quello di pompare il calcio verso l’esterno – permette anche un flusso di sodio verso l’interno, contrastando così la pompa sodio-potassio, ma, poiché le concentrazioni complessive di sodio e potassio sono molto più alte di quelle del calcio, questo effetto è relativamente poco importante. Il risultato netto dello scambiatore sodio-calcio è che nello stato di riposo, le concentrazioni intracellulari di calcio diventano molto basse.
Canali ioniciModifica

I canali ionici sono proteine integrali di membrana con un poro attraverso il quale gli ioni possono viaggiare tra lo spazio extracellulare e l’interno delle cellule. La maggior parte dei canali sono specifici (selettivi) per uno ione; per esempio, la maggior parte dei canali del potassio sono caratterizzati da un rapporto di selettività 1000:1 per il potassio rispetto al sodio, anche se gli ioni potassio e sodio hanno la stessa carica e differiscono solo leggermente nel loro raggio. Il poro del canale è tipicamente così piccolo che gli ioni devono attraversarlo in ordine di fila. I pori del canale possono essere aperti o chiusi per il passaggio degli ioni, anche se un certo numero di canali dimostrano vari livelli di subconduttanza. Quando un canale è aperto, gli ioni permeano attraverso il poro del canale lungo il gradiente di concentrazione transmembrana per quel particolare ione. La velocità del flusso ionico attraverso il canale, cioè l’ampiezza della corrente monocanale, è determinata dalla conduttanza massima del canale e dalla forza motrice elettrochimica per quello ione, che è la differenza tra il valore istantaneo del potenziale di membrana e il valore del potenziale di inversione.

Un canale può avere diversi stati (corrispondenti a diverse conformazioni della proteina), ma ognuno di questi stati è aperto o chiuso. In generale, gli stati chiusi corrispondono o a una contrazione del poro – rendendolo impraticabile per lo ione – o a una parte separata della proteina, che blocca il poro. Per esempio, il canale del sodio voltaggio-dipendente subisce l’inattivazione, in cui una parte della proteina oscilla nel poro, sigillandolo. Questa inattivazione spegne la corrente di sodio e gioca un ruolo critico nel potenziale d’azione.
I canali ionici possono essere classificati in base a come rispondono al loro ambiente. Per esempio, i canali ionici coinvolti nel potenziale d’azione sono canali sensibili alla tensione; si aprono e si chiudono in risposta alla tensione attraverso la membrana. I canali Ligand-gated formano un’altra classe importante; questi canali ionici si aprono e si chiudono in risposta al legame di una molecola di ligando, come un neurotrasmettitore. Altri canali ionici si aprono e si chiudono con forze meccaniche. Ancora altri canali ionici – come quelli dei neuroni sensoriali – si aprono e si chiudono in risposta ad altri stimoli, come la luce, la temperatura o la pressione.
Canali di perditaModifica
I canali di perdita sono il tipo più semplice di canale ionico, in quanto la loro permeabilità è più o meno costante. I tipi di canali di perdita che hanno la maggiore importanza nei neuroni sono i canali del potassio e del cloruro. Anche questi non sono perfettamente costanti nelle loro proprietà: In primo luogo, la maggior parte di essi sono voltaggio-dipendenti, nel senso che conducono meglio in una direzione che nell’altra (in altre parole, sono raddrizzatori); in secondo luogo, alcuni di essi sono in grado di essere spenti da ligandi chimici, anche se non richiedono ligandi per funzionare.
Canali a ligandiModifica
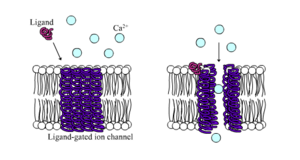
I canali ionici a ligandigated ion channels sono canali la cui permeabilità aumenta notevolmente quando un qualche tipo di ligando chimico si lega alla struttura della proteina. Le cellule animali ne contengono centinaia, se non migliaia, di tipi. Un grande sottoinsieme funziona come recettori di neurotrasmettitori – si verificano in siti postsinaptici, e il ligando chimico che li apre è rilasciato dal terminale dell’assone presinaptico. Un esempio di questo tipo è il recettore AMPA, un recettore per il neurotrasmettitore glutammato che quando viene attivato permette il passaggio di ioni sodio e potassio. Un altro esempio è il recettore GABAA, un recettore per il neurotrasmettitore GABA che quando viene attivato permette il passaggio di ioni cloruro.
I recettori neurotrasmettitori sono attivati da ligandi che appaiono nell’area extracellulare, ma ci sono altri tipi di canali ligando-gated che sono controllati da interazioni sul lato intracellulare.
Canali voltaggio-dipendenti
I canali ionici voltaggio-dipendenti, conosciuti anche come canali ionici voltaggio-dipendenti, sono canali la cui permeabilità è influenzata dal potenziale di membrana. Formano un altro gruppo molto grande, con ogni membro che ha una particolare selettività ionica e una particolare dipendenza dalla tensione. Molti sono anche dipendenti dal tempo – in altre parole, non rispondono immediatamente a un cambiamento di tensione ma solo dopo un ritardo.
Uno dei membri più importanti di questo gruppo è un tipo di canale del sodio voltaggio-gated che è alla base dei potenziali d’azione – questi sono a volte chiamati canali del sodio Hodgkin-Huxley perché sono stati inizialmente caratterizzati da Alan Lloyd Hodgkin e Andrew Huxley nei loro studi sulla fisiologia del potenziale d’azione, vincitori del premio Nobel. Il canale è chiuso al livello di tensione di riposo, ma si apre bruscamente quando la tensione supera una certa soglia, permettendo un grande afflusso di ioni sodio che produce un cambiamento molto rapido nel potenziale di membrana. Il recupero da un potenziale d’azione dipende in parte da un tipo di canale per il potassio, che è chiuso al livello di tensione di riposo, ma si apre come conseguenza del grande cambiamento di tensione prodotto durante il potenziale d’azione.
Potenziale d’inversioneModifica
Il potenziale d’inversione (o potenziale d’equilibrio) di uno ione è il valore della tensione transmembrana al quale le forze diffusionali ed elettriche si compensano, in modo che non ci sia un flusso netto di ioni attraverso la membrana. Ciò significa che la tensione transmembrana si oppone esattamente alla forza di diffusione dello ione, in modo che la corrente netta dello ione attraverso la membrana sia zero e immutabile. Il potenziale di inversione è importante perché dà la tensione che agisce sui canali permeabili a quello ione – in altre parole, dà la tensione che il gradiente di concentrazione dello ione genera quando agisce come una batteria.
Il potenziale di equilibrio di un particolare ione è solitamente designato dalla notazione Eion.Il potenziale di equilibrio per qualsiasi ione può essere calcolato usando l’equazione di Nernst. Per esempio, il potenziale di inversione per gli ioni di potassio sarà il seguente:
E e q , K + = R T z F ln o i , {\displaystyle E_{eq,K^{+}}={frac {RT}{zF}}ln {\frac {_{o}{_{i}},}

dove
- Eeq,K+ è il potenziale di equilibrio per il potassio, misurato in volt
- R è la costante universale dei gas, pari a 8,314 joule-K-1-mol-1
- T è la temperatura assoluta, misurata in kelvin (= K = gradi Celsius + 273.15)
- z è il numero di cariche elementari dello ione in questione coinvolto nella reazione
- F è la costante di Faraday, pari a 96.485 coulombs-mol-1 o J-V-1-mol-1
- o è la concentrazione extracellulare di potassio, misurata in mol-m-3 o mmol-l-1
- i è la concentrazione intracellulare di potassio
Anche se due ioni diversi hanno la stessa carica (es.e., K+ e Na+), possono ancora avere potenziali di equilibrio molto diversi, se le loro concentrazioni esterne e/o interne differiscono. Prendiamo, per esempio, i potenziali di equilibrio del potassio e del sodio nei neuroni. Il potenziale di equilibrio del potassio EK è -84 mV con 5 mM di potassio fuori e 140 mM dentro. D’altra parte, il potenziale di equilibrio del sodio, ENa, è circa +66 mV con circa 12 mM di sodio all’interno e 140 mM all’esterno.
Cambiamenti al potenziale di membrana durante lo sviluppoModifica
Il potenziale di membrana a riposo di un neurone cambia effettivamente durante lo sviluppo di un organismo. Affinché un neurone possa infine adottare la sua piena funzione adulta, il suo potenziale deve essere strettamente regolato durante lo sviluppo. Mentre un organismo progredisce nello sviluppo, il potenziale di membrana a riposo diventa più negativo. Anche le cellule gliali si differenziano e proliferano con il progredire dello sviluppo nel cervello. L’aggiunta di queste cellule gliali aumenta la capacità dell’organismo di regolare il potassio extracellulare. Il calo del potassio extracellulare può portare a una diminuzione del potenziale di membrana di 35 mV.
Eccitabilità cellulareModifica
L’eccitabilità cellulare è il cambiamento del potenziale di membrana che è necessario per le risposte cellulari in vari tessuti. L’eccitabilità cellulare è una proprietà che viene indotta durante la prima embriogenesi. L’eccitabilità di una cellula è stata anche definita come la facilità con cui una risposta può essere innescata. I potenziali di riposo e di soglia costituiscono la base dell’eccitabilità cellulare e questi processi sono fondamentali per la generazione di potenziali graduali e d’azione.
I più importanti regolatori dell’eccitabilità cellulare sono le concentrazioni di elettroliti extracellulari (cioè Na+, K+, Ca2+, Cl-, Mg2+) e le proteine associate. Proteine importanti che regolano l’eccitabilità cellulare sono i canali ionici voltaggio-gettati, i trasportatori di ioni (per esempio Na+/K+-ATPasi, trasportatori di magnesio, trasportatori acido-base), i recettori di membrana e i canali ciclico-nucleotidici iperpolarizzati. Per esempio, i canali del potassio e i recettori sensibili al calcio sono importanti regolatori dell’eccitabilità nei neuroni, nei miociti cardiaci e in molte altre cellule eccitabili come gli astrociti. Lo ione calcio è anche il secondo messaggero più importante nella segnalazione delle cellule eccitabili. L’attivazione dei recettori sinaptici avvia cambiamenti di lunga durata nell’eccitabilità neuronale. Anche gli ormoni tiroidei, surrenali e altri regolano l’eccitabilità cellulare.
Molti tipi di cellule sono considerati avere una membrana eccitabile. Le cellule eccitabili sono neuroni, miociti (cardiaci, scheletrici, lisci), cellule vascolari endoteliali, cellule juxtaglomerulari, cellule interstiziali di Cajal, molti tipi di cellule epiteliali (es. cellule beta, cellule alfa, cellule delta, cellule enteroendocrine), cellule gliali (per esempio astrociti), cellule meccanorecettrici (per esempio cellule ciliate e cellule di Merkel), cellule chemorecettrici (per esempio cellule del glomo, recettori del gusto), alcune cellule vegetali e forse cellule immunitarie. Gli astrociti mostrano una forma di eccitabilità non elettrica basata su variazioni intracellulari di calcio legate all’espressione di diversi recettori attraverso i quali possono rilevare il segnale sinaptico. Nei neuroni, ci sono diverse proprietà di membrana in alcune porzioni della cellula, per esempio, l’eccitabilità dendritica conferisce ai neuroni la capacità di rilevare la coincidenza di input spazialmente separati.
Circuito equivalenteModifica
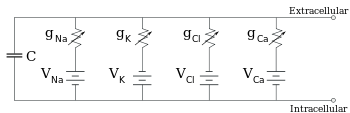
Gli elettrofisiologi modellano gli effetti delle differenze di concentrazione ionica, dei canali ionici e della capacità di membrana in termini di un circuito equivalente, che ha lo scopo di rappresentare le proprietà elettriche di una piccola patch di membrana. Il circuito equivalente consiste in un condensatore in parallelo con quattro percorsi che consistono ciascuno in una batteria in serie con una conduttanza variabile. La capacità è determinata dalle proprietà del bilayer lipidico, ed è presa per essere fissa. Ognuno dei quattro percorsi paralleli proviene da uno dei principali ioni, sodio, potassio, cloruro e calcio. La tensione di ogni via ionica è determinata dalle concentrazioni dello ione su ogni lato della membrana; vedi la sezione Potenziale d’inversione sopra. La conduttanza di ogni via ionica in qualsiasi momento è determinata dagli stati di tutti i canali ionici che sono potenzialmente permeabili a quello ione, compresi i canali di perdita, i canali ligando-gated e i canali ionici voltage-gated.
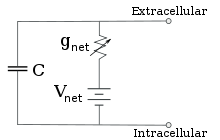
Per concentrazioni fisse di ioni e valori fissi della conduttanza dei canali ionici, il circuito equivalente può essere ulteriormente ridotto, utilizzando l’equazione di Goldman come descritto di seguito, a un circuito contenente una capacità in parallelo con una batteria e la conduttanza. In termini elettrici, questo è un tipo di circuito RC (circuito resistenza-capacità), e le sue proprietà elettriche sono molto semplici. A partire da qualsiasi stato iniziale, la corrente che scorre attraverso la conduttanza o la capacità decade con un corso di tempo esponenziale, con una costante di tempo di τ = RC, dove C è la capacità del patch di membrana, e R = 1/gnet è la resistenza netta. Per situazioni realistiche, la costante di tempo si trova di solito nell’intervallo 1-100 millisecondi. Nella maggior parte dei casi, i cambiamenti nella conduttanza dei canali ionici si verificano su una scala temporale più veloce, quindi un circuito RC non è una buona approssimazione; tuttavia, l’equazione differenziale utilizzata per modellare un patch di membrana è comunemente una versione modificata dell’equazione del circuito RC.