Membranpotential
Das Membranpotential in einer Zelle ergibt sich letztlich aus zwei Faktoren: elektrische Kraft und Diffusion. Die elektrische Kraft ergibt sich aus der gegenseitigen Anziehung zwischen Teilchen mit entgegengesetzten elektrischen Ladungen (positiv und negativ) und der gegenseitigen Abstoßung zwischen Teilchen mit der gleichen Art von Ladung (beide positiv oder beide negativ). Diffusion entsteht durch die statistische Tendenz von Teilchen, sich von Regionen, in denen sie hoch konzentriert sind, in Regionen mit geringer Konzentration umzuverteilen.
VoltageEdit
Spannung, gleichbedeutend mit elektrischer Potentialdifferenz, ist die Fähigkeit, einen elektrischen Strom über einen Widerstand zu treiben. In der Tat ist die einfachste Definition einer Spannung durch das Ohmsche Gesetz gegeben: V=IR, wobei V die Spannung, I der Strom und R der Widerstand ist. Wenn eine Spannungsquelle, wie z. B. eine Batterie, in einem elektrischen Stromkreis platziert wird, ist die Strommenge, die sie über den verfügbaren Widerstand treibt, umso größer, je höher die Spannung der Quelle ist. Die funktionelle Bedeutung von Spannung liegt nur in Potentialdifferenzen zwischen zwei Punkten in einem Stromkreis. Die Vorstellung von einer Spannung an einem einzelnen Punkt ist bedeutungslos. In der Elektronik ist es üblich, einem willkürlich gewählten Element der Schaltung eine Spannung von Null zuzuordnen und dann Spannungen für andere Elemente zuzuordnen, die relativ zu diesem Nullpunkt gemessen werden. Es ist nicht von Bedeutung, welches Element als Nullpunkt gewählt wird – die Funktion einer Schaltung hängt nur von den Unterschieden ab, nicht von den Spannungen an sich. In den meisten Fällen und nach Konvention wird der Nullpunkt jedoch dem Teil eines Stromkreises zugeordnet, der mit der Masse in Kontakt ist.
Das gleiche Prinzip gilt für die Spannung in der Zellbiologie. In elektrisch aktivem Gewebe kann die Potentialdifferenz zwischen zwei beliebigen Punkten gemessen werden, indem man an jedem Punkt eine Elektrode anbringt, z. B. eine innerhalb und eine außerhalb der Zelle, und beide Elektroden mit den Leitungen eines im Wesentlichen spezialisierten Voltmeters verbindet. Das Vorzeichen der Potentialdifferenz zwischen Außen- und Innenseite wird durch das Potential der Innenseite relativ zum äußeren Nullpunkt bestimmt.
Mathematisch beginnt die Definition der Spannung mit dem Konzept des elektrischen Feldes E, einem Vektorfeld, das jedem Punkt im Raum eine Größe und Richtung zuordnet. In vielen Situationen ist das elektrische Feld ein konservatives Feld, was bedeutet, dass es als Gradient einer Skalarfunktion V ausgedrückt werden kann, also E = -∇V. Dieses Skalarfeld V wird als Spannungsverteilung bezeichnet. Beachten Sie, dass die Definition eine beliebige Integrationskonstante zulässt – deshalb sind absolute Werte der Spannung nicht sinnvoll. Im Allgemeinen können elektrische Felder nur dann als konservativ behandelt werden, wenn magnetische Felder sie nicht signifikant beeinflussen, aber diese Bedingung trifft in der Regel gut auf biologisches Gewebe zu.
Da das elektrische Feld der Gradient der Spannungsverteilung ist, implizieren schnelle Spannungsänderungen innerhalb einer kleinen Region ein starkes elektrisches Feld; umgekehrt müssen, wenn die Spannung über eine große Region annähernd gleich bleibt, die elektrischen Felder in dieser Region schwach sein. Ein starkes elektrisches Feld, gleichbedeutend mit einem starken Spannungsgradienten, impliziert, dass eine starke Kraft auf alle geladenen Teilchen ausgeübt wird, die sich innerhalb der Region befinden.
Ionen und die Kräfte, die ihre Bewegung antreibenBearbeiten

Elektrische Signale in biologischen Organismen werden im Allgemeinen von Ionen angetrieben. Die wichtigsten Kationen für das Aktionspotential sind Natrium (Na+) und Kalium (K+). Beides sind monovalente Kationen, die eine einzelne positive Ladung tragen. An Aktionspotentialen kann auch Calcium (Ca2+) beteiligt sein, das als zweiwertiges Kation eine doppelte positive Ladung trägt. Das Chlorid-Anion (Cl-) spielt eine große Rolle in den Aktionspotentialen einiger Algen, spielt aber eine vernachlässigbare Rolle in den Aktionspotentialen der meisten Tiere.
Ionen durchqueren die Zellmembran unter zwei Einflüssen: Diffusion und elektrische Felder. Ein einfaches Beispiel, bei dem zwei Lösungen – A und B – durch eine poröse Barriere getrennt sind, zeigt, dass die Diffusion dafür sorgt, dass sie sich schließlich zu gleichen Lösungen vermischen. Diese Vermischung erfolgt aufgrund des Unterschieds in ihren Konzentrationen. Der Bereich mit der hohen Konzentration diffundiert in Richtung des Bereichs mit der niedrigen Konzentration aus. Um das Beispiel zu erweitern, soll Lösung A 30 Natriumionen und 30 Chloridionen enthalten. Lösung B habe nur 20 Natriumionen und 20 Chloridionen. Angenommen, die Barriere lässt beide Ionenarten durch, dann wird ein stationärer Zustand erreicht, bei dem beide Lösungen 25 Natriumionen und 25 Chloridionen aufweisen. Wenn die poröse Barriere jedoch selektiv ist, welche Ionen durchgelassen werden, dann wird die Diffusion allein nicht die resultierende Lösung bestimmen. Kehren wir zum vorherigen Beispiel zurück und konstruieren wir nun eine Barriere, die nur für Natriumionen durchlässig ist. Jetzt wird nur Natrium erlaubt, durch die Barriere von seiner höheren Konzentration in Lösung A zu der niedrigeren Konzentration in Lösung B zu diffundieren. Dies führt zu einer größeren Anhäufung von Natriumionen als Chloridionen in Lösung B und einer geringeren Anzahl von Natriumionen als Chloridionen in Lösung A.
Das bedeutet, dass es eine positive Nettoladung in Lösung B durch die höhere Konzentration von positiv geladenen Natriumionen als negativ geladenen Chloridionen gibt. Ebenso gibt es eine negative Nettoladung in Lösung A durch die höhere Konzentration an negativen Chloridionen als an positiven Natriumionen. Da sich entgegengesetzte Ladungen anziehen und gleichartige Ladungen abstoßen, werden die Ionen nun auch von elektrischen Feldern sowie von Diffusionskräften beeinflusst. Daher werden die positiven Natriumionen weniger wahrscheinlich in die nun positivere B-Lösung wandern und in der nun negativeren A-Lösung verbleiben. Der Punkt, an dem die Kräfte der elektrischen Felder der Kraft aufgrund der Diffusion vollständig entgegenwirken, wird als Gleichgewichtspotential bezeichnet. An diesem Punkt ist der Nettostrom des jeweiligen Ions (in diesem Fall Natrium) gleich Null.
PlasmamembranenBearbeiten

Jede Zelle ist von einer Plasmamembran umschlossen, die die Struktur einer Lipiddoppelschicht hat, in die viele Arten von großen Molekülen eingebettet sind. Da sie aus Lipidmolekülen besteht, hat die Plasmamembran intrinsisch einen hohen elektrischen Widerstand, also eine geringe intrinsische Permeabilität für Ionen. Einige der in der Membran eingebetteten Moleküle sind jedoch in der Lage, Ionen entweder aktiv von einer Seite der Membran zur anderen zu transportieren oder Kanäle bereitzustellen, durch die sie sich bewegen können.
In der elektrischen Terminologie funktioniert die Plasmamembran als eine Kombination aus Widerstand und Kondensator. Der Widerstand ergibt sich aus der Tatsache, dass die Membran die Bewegung von Ladungen durch sie behindert. Die Kapazität ergibt sich aus der Tatsache, dass die Lipiddoppelschicht so dünn ist, dass eine Ansammlung geladener Teilchen auf einer Seite eine elektrische Kraft erzeugt, die entgegengesetzt geladene Teilchen auf die andere Seite zieht. Die Kapazität der Membran ist relativ unbeeinflusst von den darin eingebetteten Molekülen, so dass sie einen mehr oder weniger unveränderlichen Wert hat, der auf etwa 2 μF/cm2 geschätzt wird (die Gesamtkapazität eines Membranflecks ist proportional zu seiner Fläche). Die Leitfähigkeit einer reinen Lipiddoppelschicht ist dagegen so gering, dass sie in biologischen Situationen immer von der Leitfähigkeit alternativer Wege, die durch eingebettete Moleküle bereitgestellt werden, dominiert wird. Die Kapazität der Membran ist also mehr oder weniger fest, der Widerstand jedoch stark variabel.
Die Dicke einer Plasmamembran wird auf etwa 7-8 Nanometer geschätzt. Weil die Membran so dünn ist, braucht es keine sehr große Transmembranspannung, um in ihr ein starkes elektrisches Feld zu erzeugen. Typische Membranpotentiale in tierischen Zellen liegen in der Größenordnung von 100 Millivolt (d.h. ein Zehntel Volt), aber Berechnungen zeigen, dass dies ein elektrisches Feld erzeugt, das nahe am Maximum liegt, das die Membran aushalten kann – es wurde berechnet, dass eine Spannungsdifferenz, die viel größer als 200 Millivolt ist, einen dielektrischen Durchschlag, d.h. einen Lichtbogen über die Membran, verursachen könnte.
Erleichterte Diffusion und TransportBearbeiten

Der Widerstand einer reinen Lipiddoppelschicht gegenüber dem Durchtritt von Ionen ist sehr hoch, aber in die Membran eingebettete Strukturen können die Ionenbewegung stark verbessern, entweder aktiv oder passiv, über Mechanismen, die als erleichterter Transport und erleichterte Diffusion bezeichnet werden. Die beiden Strukturtypen, die die größte Rolle spielen, sind Ionenkanäle und Ionenpumpen, die beide in der Regel aus Ansammlungen von Proteinmolekülen bestehen. Ionenkanäle bieten Durchgänge, durch die sich Ionen bewegen können. In den meisten Fällen ist ein Ionenkanal nur für bestimmte Arten von Ionen durchlässig (z. B. Natrium und Kalium, aber nicht Chlorid oder Kalzium), und manchmal variiert die Durchlässigkeit in Abhängigkeit von der Richtung der Ionenbewegung. Ionenpumpen, auch Ionentransporter oder Carrierproteine genannt, transportieren aktiv bestimmte Ionenarten von einer Seite der Membran zur anderen, wobei sie manchmal Energie aus Stoffwechselprozessen dafür verwenden.
IonenpumpenBearbeiten
Hauptartikel: Ionentransporter und Aktiver Transport
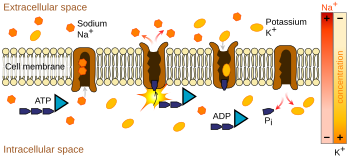
Ionenpumpen sind integrale Membranproteine, die einen aktiven Transport durchführen, d. h. sie nutzen zelluläre Energie (ATP), um die Ionen gegen ihren Konzentrationsgradienten zu „pumpen“. Solche Ionenpumpen nehmen auf der einen Seite der Membran Ionen auf (verringern dort die Konzentration) und geben sie auf der anderen Seite wieder ab (erhöhen dort die Konzentration).
Die für das Aktionspotential relevanteste Ionenpumpe ist die Natrium-Kalium-Pumpe, die drei Natrium-Ionen aus der Zelle heraus und zwei Kalium-Ionen hinein transportiert. Dies hat zur Folge, dass die Konzentration der Kalium-Ionen K+ im Inneren des Neurons etwa 20-mal größer ist als die Konzentration außerhalb, während die Natrium-Konzentration außen etwa neunmal größer ist als innen. In ähnlicher Weise haben auch andere Ionen unterschiedliche Konzentrationen innerhalb und außerhalb des Neurons, wie z. B. Kalzium, Chlorid und Magnesium.
Wären die Anzahlen jeder Ionenart gleich, wäre die Natrium-Kalium-Pumpe elektrisch neutral, aber wegen des Drei-für-Zwei-Austauschs bewirkt sie bei jedem Zyklus eine Nettobewegung einer positiven Ladung von intrazellulär nach extrazellulär und trägt damit zu einer positiven Spannungsdifferenz bei. Die Pumpe hat drei Effekte: (1) sie sorgt dafür, dass die Natriumkonzentration im Extrazellulärraum hoch und im Intrazellulärraum niedrig ist; (2) sie sorgt dafür, dass die Kaliumkonzentration im Intrazellulärraum hoch und im Extrazellulärraum niedrig ist; (3) sie verleiht dem Intrazellulärraum eine negative Spannung in Bezug auf den Extrazellulärraum.
Die Natrium-Kalium-Pumpe ist relativ langsam im Betrieb. Wenn eine Zelle mit überall gleichen Konzentrationen von Natrium und Kalium initialisiert würde, würde es Stunden dauern, bis die Pumpe ein Gleichgewicht herstellt. Die Pumpe arbeitet konstant, wird aber zunehmend ineffizienter, wenn die Konzentrationen von Natrium und Kalium, die für das Pumpen zur Verfügung stehen, reduziert werden.
Ionenpumpen beeinflussen das Aktionspotential nur, indem sie das relative Verhältnis von intrazellulären und extrazellulären Ionenkonzentrationen festlegen. Das Aktionspotenzial beinhaltet hauptsächlich das Öffnen und Schließen von Ionenkanälen, nicht von Ionenpumpen. Wenn die Ionenpumpen ausgeschaltet werden, indem man ihnen die Energiequelle entzieht oder einen Inhibitor wie z. B. Ouabain hinzufügt, kann das Axon immer noch Hunderttausende von Aktionspotenzialen abfeuern, bevor ihre Amplituden signifikant abzufallen beginnen. Insbesondere spielen Ionenpumpen bei der Repolarisation der Membran nach einem Aktionspotential keine wesentliche Rolle.
Eine weitere funktionell wichtige Ionenpumpe ist der Natrium-Calcium-Austauscher. Diese Pumpe arbeitet konzeptionell ähnlich wie die Natrium-Kalium-Pumpe, nur dass sie in jedem Zyklus drei Na+ aus dem Extrazellulärraum gegen ein Ca++ aus dem Intrazellulärraum austauscht. Da der Nettoladungsfluss nach innen gerichtet ist, läuft diese Pumpe gewissermaßen „bergab“ und benötigt daher außer der Membranspannung keine Energiequelle. Ihr wichtigster Effekt ist es, Kalzium nach außen zu pumpen – sie ermöglicht auch einen Einstrom von Natrium und wirkt damit der Natrium-Kalium-Pumpe entgegen, aber da die Natrium- und Kalium-Konzentrationen insgesamt viel höher sind als die Kalzium-Konzentrationen, ist dieser Effekt relativ unbedeutend. Das Nettoergebnis des Natrium-Kalzium-Austauschers ist, dass im Ruhezustand die intrazellulären Kalziumkonzentrationen sehr niedrig werden.
IonenkanäleBearbeiten

Ionenkanäle sind integrale Membranproteine mit einer Pore, durch die Ionen zwischen Extrazellulärraum und Zellinnerem wandern können. Die meisten Kanäle sind spezifisch (selektiv) für ein Ion; zum Beispiel haben die meisten Kaliumkanäle ein Selektivitätsverhältnis von 1000:1 für Kalium gegenüber Natrium, obwohl Kalium- und Natriumionen die gleiche Ladung haben und sich nur geringfügig in ihrem Radius unterscheiden. Die Kanalpore ist typischerweise so klein, dass die Ionen sie in einreihiger Reihenfolge passieren müssen. Die Kanalporen können für den Ionendurchgang entweder offen oder geschlossen sein, wobei eine Reihe von Kanälen verschiedene Unterleitfähigkeiten aufweisen. Wenn ein Kanal offen ist, permeieren Ionen durch die Kanalpore entlang des Transmembrankonzentrationsgradienten für das jeweilige Ion. Die Geschwindigkeit des Ionenflusses durch den Kanal, d. h. die Stromamplitude des Einzelkanals, wird durch die maximale Kanalleitfähigkeit und die elektrochemische Antriebskraft für dieses Ion bestimmt, die die Differenz zwischen dem Momentanwert des Membranpotenzials und dem Wert des Umkehrpotenzials ist.

Ein Kanal kann mehrere verschiedene Zustände haben (entsprechend verschiedenen Konformationen des Proteins), aber jeder dieser Zustände ist entweder offen oder geschlossen. Im Allgemeinen entsprechen geschlossene Zustände entweder einer Verengung der Pore – wodurch sie für das Ion unpassierbar wird – oder einem separaten Teil des Proteins, der die Pore verschließt. Der spannungsabhängige Natriumkanal beispielsweise erfährt eine Inaktivierung, bei der ein Teil des Proteins in die Pore einschwenkt und diese verschließt. Diese Inaktivierung schaltet den Natriumstrom ab und spielt eine entscheidende Rolle für das Aktionspotential.
Ionenkanäle können danach klassifiziert werden, wie sie auf ihre Umgebung reagieren. Die Ionenkanäle, die am Aktionspotenzial beteiligt sind, sind zum Beispiel spannungsempfindliche Kanäle; sie öffnen und schließen sich in Abhängigkeit von der Spannung an der Membran. Ligandengesteuerte Kanäle bilden eine weitere wichtige Klasse; diese Ionenkanäle öffnen und schließen sich als Reaktion auf die Bindung eines Ligandenmoleküls, z. B. eines Neurotransmitters. Andere Ionenkanäle öffnen und schließen sich durch mechanische Kräfte. Wieder andere Ionenkanäle – wie die von sensorischen Neuronen – öffnen und schließen sich als Reaktion auf andere Reize, wie Licht, Temperatur oder Druck.
LeckagekanäleBearbeiten
Leckagekanäle sind die einfachste Art von Ionenkanälen, da ihre Permeabilität mehr oder weniger konstant ist. Die Typen von Leckagekanälen, die in Neuronen die größte Bedeutung haben, sind Kalium- und Chloridkanäle. Auch diese sind in ihren Eigenschaften nicht vollkommen konstant: Erstens sind die meisten von ihnen spannungsabhängig in dem Sinne, dass sie in eine Richtung besser leiten als in die andere (sie sind also Gleichrichter); zweitens sind einige von ihnen in der Lage, durch chemische Liganden abgeschaltet zu werden, obwohl sie keine Liganden benötigen, um zu funktionieren.
Ligandengesteuerte KanäleBearbeiten
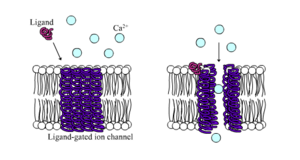
Liganden-gated ion channels sind Kanäle, deren Permeabilität stark erhöht ist, wenn eine Art chemischer Ligand an die Proteinstruktur bindet. Tierische Zellen enthalten Hunderte, wenn nicht Tausende von Typen dieser Kanäle. Eine große Untergruppe fungiert als Neurotransmitter-Rezeptoren – sie kommen an postsynaptischen Stellen vor, und der chemische Ligand, der sie schaltet, wird vom präsynaptischen Axonterminal freigesetzt. Ein Beispiel für diesen Typ ist der AMPA-Rezeptor, ein Rezeptor für den Neurotransmitter Glutamat, der, wenn er aktiviert wird, den Durchgang von Natrium- und Kaliumionen ermöglicht. Ein anderes Beispiel ist der GABAA-Rezeptor, ein Rezeptor für den Neurotransmitter GABA, der, wenn er aktiviert wird, den Durchgang von Chlorid-Ionen erlaubt.
Neurotransmitter-Rezeptoren werden durch Liganden aktiviert, die im extrazellulären Bereich auftreten, aber es gibt auch andere Arten von ligandengesteuerten Kanälen, die durch Wechselwirkungen auf der intrazellulären Seite gesteuert werden.
Spannungsabhängige Kanäle
Spannungsabhängige Ionenkanäle, auch als spannungsabhängige Ionenkanäle bezeichnet, sind Kanäle, deren Permeabilität durch das Membranpotential beeinflusst wird. Sie bilden eine weitere sehr große Gruppe, wobei jedes Mitglied eine bestimmte Ionenselektivität und eine bestimmte Spannungsabhängigkeit aufweist. Viele sind auch zeitabhängig, das heißt, sie reagieren nicht sofort auf eine Spannungsänderung, sondern erst mit einer Verzögerung.
Einer der wichtigsten Vertreter dieser Gruppe ist ein Typ von spannungsabhängigen Natriumkanälen, der den Aktionspotentialen zugrunde liegt – sie werden manchmal auch Hodgkin-Huxley-Natriumkanäle genannt, weil sie ursprünglich von Alan Lloyd Hodgkin und Andrew Huxley in ihren mit dem Nobelpreis ausgezeichneten Studien zur Physiologie des Aktionspotentials charakterisiert wurden. Der Kanal ist bei der Ruhespannung geschlossen, öffnet sich aber schlagartig, wenn die Spannung einen bestimmten Schwellenwert überschreitet, und ermöglicht so einen großen Einstrom von Natriumionen, der eine sehr schnelle Änderung des Membranpotentials bewirkt. Die Erholung von einem Aktionspotential hängt teilweise von einer Art spannungsgesteuertem Kaliumkanal ab, der auf dem Niveau der Ruhespannung geschlossen ist, sich aber als Folge der großen Spannungsänderung, die während des Aktionspotentials erzeugt wird, öffnet.
UmkehrpotentialBearbeiten
Das Umkehrpotential (oder Gleichgewichtspotential) eines Ions ist der Wert der Transmembranspannung, bei dem sich diffusive und elektrische Kräfte ausgleichen, so dass kein Netto-Ionenfluss über die Membran stattfindet. Das bedeutet, dass die Transmembranspannung der Diffusionskraft des Ions genau entgegengesetzt ist, so dass der Nettostrom des Ions durch die Membran null und unveränderlich ist. Das Umkehrpotential ist wichtig, weil es die Spannung angibt, die auf Kanäle wirkt, die für dieses Ion durchlässig sind – mit anderen Worten, es gibt die Spannung an, die das Ionenkonzentrationsgefälle erzeugt, wenn es als Batterie wirkt.
Das Gleichgewichtspotential eines bestimmten Ions wird üblicherweise mit der Schreibweise Eion bezeichnet.
Das Gleichgewichtspotential für jedes Ion kann mit Hilfe der Nernst-Gleichung berechnet werden. Zum Beispiel lautet das Umkehrpotential für Kalium-Ionen wie folgt:
E e q , K + = R T z F ln o i , {\displaystyle E_{eq,K^{+}}={\frac {RT}{zF}}\ln {\frac {_{o}}{_{i}},}

wobei
- Eeq,K+ ist das Gleichgewichtspotential für Kalium, gemessen in Volt
- R ist die universelle Gaskonstante, gleich 8,314 Joule-K-1-mol-1
- T ist die absolute Temperatur, gemessen in Kelvin (= K = Grad Celsius + 273.15)
- z ist die Anzahl der Elementarladungen des betreffenden Ions, die an der Reaktion beteiligt sind
- F ist die Faraday-Konstante, gleich 96,485 Coulomb-mol-1 oder J-V-1-mol-1
- o ist die extrazelluläre Konzentration von Kalium, gemessen in mol-m-3 oder mmol-l-1
- i ist die intrazelluläre Konzentration von Kalium
Auch wenn zwei verschiedene Ionen die gleiche Ladung haben (d. h.e., K+ und Na+), können sie dennoch sehr unterschiedliche Gleichgewichtspotentiale haben, sofern sich ihre äußeren und/oder inneren Konzentrationen unterscheiden. Nehmen wir zum Beispiel die Gleichgewichtspotentiale von Kalium und Natrium in Neuronen. Das Kalium-Gleichgewichtspotential EK beträgt -84 mV bei 5 mM Kalium außen und 140 mM innen. Auf der anderen Seite liegt das Natrium-Gleichgewichtspotential ENa bei etwa +66 mV mit etwa 12 mM Natrium innen und 140 mM außen.
Änderungen des Membranpotentials während der Entwicklung
Das Ruhemembranpotential eines Neurons ändert sich tatsächlich während der Entwicklung eines Organismus. Damit ein Neuron schließlich seine volle Funktion als Erwachsener übernehmen kann, muss sein Potenzial während der Entwicklung streng reguliert werden. Im Laufe der Entwicklung eines Organismus wird das Ruhemembranpotenzial negativer. Auch Gliazellen differenzieren und proliferieren im Laufe der Entwicklung im Gehirn. Das Hinzukommen dieser Gliazellen erhöht die Fähigkeit des Organismus, das extrazelluläre Kalium zu regulieren. Der Abfall des extrazellulären Kaliums kann zu einer Abnahme des Membranpotentials um 35 mV führen.
Zell-ErregbarkeitBearbeiten
Zell-Erregbarkeit ist die Änderung des Membranpotentials, die für zelluläre Reaktionen in verschiedenen Geweben notwendig ist. Die Erregbarkeit einer Zelle ist eine Eigenschaft, die während der frühen Embriogenese induziert wird. Die Erregbarkeit einer Zelle wurde auch als die Leichtigkeit definiert, mit der eine Reaktion ausgelöst werden kann. Die Ruhe- und Schwellenpotentiale bilden die Grundlage der Zellerregbarkeit und diese Prozesse sind grundlegend für die Erzeugung von Gradienten- und Aktionspotentialen.
Die wichtigsten Regulatoren der Zellerregbarkeit sind die extrazellulären Elektrolytkonzentrationen (d.h. Na+, K+, Ca2+, Cl-, Mg2+) und assoziierte Proteine. Wichtige Proteine, die die Erregbarkeit der Zelle regulieren, sind spannungsabhängige Ionenkanäle, Ionentransporter (z. B. Na+/K+-ATPase, Magnesiumtransporter, Säure-Basen-Transporter), Membranrezeptoren und hyperpolarisationsaktivierte zyklische Nukleotid-abhängige Kanäle. Zum Beispiel sind Kaliumkanäle und Calcium-sensing Rezeptoren wichtige Regulatoren der Erregbarkeit in Neuronen, Herzmuskelzellen und vielen anderen erregbaren Zellen wie Astrozyten. Das Kalzium-Ion ist auch der wichtigste Second Messenger bei der Signalübertragung in erregbaren Zellen. Die Aktivierung synaptischer Rezeptoren löst lang anhaltende Veränderungen der neuronalen Erregbarkeit aus. Auch Schilddrüsen-, Nebennieren- und andere Hormone regulieren die Erregbarkeit von Zellen.
Viele Zelltypen gelten als erregbar. Erregbare Zellen sind Neuronen, Myozyten (kardial, skelettal, glatt), vaskuläre Endothelzellen, juxtaglomeruläre Zellen, interstitielle Zellen von Cajal, viele Arten von Epithelzellen (z.B.. Betazellen, Alphazellen, Deltazellen, enteroendokrine Zellen), Gliazellen (z. B. Astrozyten), Mechanorezeptorzellen (z. B. Haarzellen und Merkelzellen), Chemorezeptorzellen (z. B. Glomuszellen, Geschmacksrezeptoren), einige Pflanzenzellen und möglicherweise Immunzellen. Astrozyten weisen eine Form der nicht-elektrischen Erregbarkeit auf, die auf intrazellulären Kalziumschwankungen basiert und mit der Expression verschiedener Rezeptoren zusammenhängt, über die sie das synaptische Signal erkennen können. In Neuronen gibt es unterschiedliche Membraneigenschaften in einigen Teilen der Zelle, zum Beispiel verleiht die dendritische Erregbarkeit Neuronen die Fähigkeit zur Koinzidenzerkennung von räumlich getrennten Eingängen.
ÄquivalenzschaltungBearbeiten
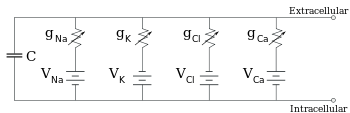
Elektrophysiologen modellieren die Effekte von Ionenkonzentrationsunterschieden, Ionenkanälen und Membrankapazität in Form eines Ersatzschaltbildes, das die elektrischen Eigenschaften eines kleinen Membranflecks darstellen soll. Das Ersatzschaltbild besteht aus einem Kondensator in Parallelschaltung mit vier Bahnen, die jeweils aus einer Batterie in Reihe mit einem variablen Leitwert bestehen. Die Kapazität wird durch die Eigenschaften der Lipiddoppelschicht bestimmt und wird als fest angenommen. Jeder der vier parallelen Pfade stammt von einem der Hauptionen, Natrium, Kalium, Chlorid und Calcium. Die Spannung jedes Ionenwegs wird durch die Konzentrationen des Ions auf jeder Seite der Membran bestimmt; siehe den Abschnitt Umkehrpotential oben. Die Leitfähigkeit jedes Ionenweges zu einem beliebigen Zeitpunkt wird durch die Zustände aller Ionenkanäle bestimmt, die potenziell für dieses Ion durchlässig sind, einschließlich Leckagekanäle, ligandengesteuerte Kanäle und spannungsgesteuerte Ionenkanäle.
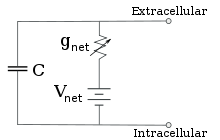
Reduzierte Schaltung, die durch Kombination der ionenspezifischen Pfade unter Verwendung der Goldman-Gleichung erhalten wird
Für feste Ionenkonzentrationen und feste Werte der Ionenkanalleitfähigkeit, kann das Ersatzschaltbild mit Hilfe der Goldman-Gleichung, wie unten beschrieben, weiter reduziert werden auf eine Schaltung, die eine Kapazität parallel zu einer Batterie und einem Leitwert enthält. Elektrisch gesehen ist dies eine Art RC-Schaltung (Widerstand-Kapazität-Schaltung), deren elektrische Eigenschaften sehr einfach sind. Ausgehend von einem beliebigen Anfangszustand fällt der Strom, der entweder über den Leitwert oder die Kapazität fließt, mit einem exponentiellen Zeitverlauf ab, mit einer Zeitkonstante von τ = RC, wobei C die Kapazität des Membranflecks und R = 1/gnet der Netzwiderstand ist. Für realistische Situationen liegt die Zeitkonstante meist im Bereich von 1-100 Millisekunden. In den meisten Fällen treten Änderungen in der Leitfähigkeit von Ionenkanälen auf einer schnelleren Zeitskala auf, so dass eine RC-Schaltung keine gute Näherung ist; die Differentialgleichung, die zur Modellierung eines Membranflecks verwendet wird, ist jedoch üblicherweise eine modifizierte Version der RC-Schaltungsgleichung.