Atlas van de planten- en dierenhistologie
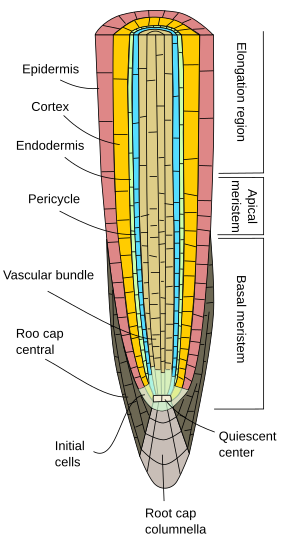
In eerste instantie groeien wortels in de lengte door de activiteit van het wortelapicale meristeem, dat wordt beschermd door de wortelkap. Proliferatie en differentiatie van de cellen van het wortelapicale meristeem geven cellen die eerst worden georganiseerd als de zogenaamde primaire wortel, dat wil zeggen dat deze primaire groei vertoont. Het is een minder complexe organisatie dan de stengel, omdat knopen, internodiën en bladeren ontbreken.
Gelijk voor welke plantensoort, vertonen primaire wortels een epidermis, of rhyzodermis, gewoonlijk uniseriaat (één dikke cellaag), zonder huidmondjes, en met wortelharen in het rijpingsgebied, net na het strekkingsgebied (figuren 1, 2, en 3). In het algemeen vertonen de epidermale cellen in de buurt van de wortelpunt een dunne cuticula, die het binnendringen van water en minerale zouten vergemakkelijkt. Bij sommige soorten zoals xerofyten en bij wortels die dicht bij het bodemoppervlak staan, bevindt zich onder de epidermis een laag cellen die hypodermis wordt genoemd. De cellen van de hypodermis vormen een dunne laag cellen met verkurkte celwanden. De hypodermis kan zich verder specialiseren en wordt de exodermis, die bij sommige angiospermen wordt aangetroffen, en werkt als een tweede barrière om de vrije verspreiding van stoffen tussen de wortel en de bodem te voorkomen.

Wortelharen nemen water en minerale zouten op. Het zijn langgerekte epidermale cellen die in het rijpingsgebied worden aangetroffen en die het absorptieoppervlak in contact met de externe omgeving vergroten, en daarmee het absorptievermogen. Wortelharen worden voortdurend gevormd en verdwijnen tijdens de groei van de wortel, omdat de rijpingsregio de groei van de wortelpunt op een min of meer constante afstand volgt. Het aantal wortelharen bedraagt ongeveer 20 tot 500 per cm2 in de wortels van bomen, en ongeveer 25000 per cm2 in de winterrogge. Het aantal kan ook variëren afhankelijk van de omgevingsomstandigheden.
Er zijn drie wortelhaarorganisatiepatronen (figuur 4). De dichtheid van de wortelharen is afhankelijk van de milieuomstandigheden. Bijvoorbeeld, in fosfaatarme bodems is het aantal wortelharen hoger om het totale wortelopnameoppervlak te vergroten. Veel symbiotische organismen, zoals stikstofbindende bacteriën, zijn verbonden met wortelharen.


De wortelcortex bevindt zich onder de epidermis, of hypodermis. In wortels is de cortex dik of zeer dik (veel meer dan in stengels), en hij bestaat meestal uit parenchymcellen die gespecialiseerd zijn in opslag, hoewel hij in luchtwortels fotosynthetisch parenchym kan zijn, en in waterwortels luchtig parenchym. Er zijn veel lege ruimten tussen de corticale parenchymcellen, zodat water kan worden geleid (apoplastische weg) naar de vasculaire bundel. Bovendien zijn de parenchymcellen onderling verbonden door plasmodesmata (symplastische weg) die ervoor zorgen dat mineralen en water van cel naar cel kunnen gaan.

Een onderscheidend kenmerk van de primaire wortelgroei is de endodermis, de binnenste laag van de cortex. De endodermis is evolutionair geconserveerd van varens tot angiospermen. Het is een ééncellige dikke laag van dicht opeengepakte cellen waarvan de celwanden gedeeltelijk met suberine zijn geïmpregneerd en zo een verdikking vormen, de zogenaamde Casparian strepen. De celverdichting en de ondoordringbaarheid van de kaspische strepen zorgen ervoor dat water en opgeloste stoffen de endodermis kunnen passeren via het cytoplasma van de endodermis cellen. Vandaar dat de endodermis een barrière vormt tegen vrije diffusie die de stoffen controleert die van de grond naar de vaatbundels gaan. In de wortelgebieden met secundaire groei zijn zowel endodermis als cortex verloren gegaan (zie volgende bladzijde).
Casparische strepen zijn primaire wandimpregnaties die de endodermale cellen omgeven als een gordel die in de lengterichting is gerangschikt. Zij zijn geen secundaire wand. De strepen zijn doorlopend tussen naburige cellen door de middenlamel. In 3D-aanzicht vormen ze als een cilindrisch visnet, waarbij de koorden de Casparian strepen zijn en de gaten de endodermis cellen. Casparian strepen bevatten lignine, maar geen suberine. In de celwand van de endodemiscellen is echter wel suberine te vinden nadat de Casparian strepen zijn gevormd. Suberine wordt als lagen over het celwandoppervlak gelegd, met een verschillende dikte naargelang de lokalisatie van de cel. De endodemiscellen die dichter bij de xyleempolen worden aangetroffen, synthetiseren bijvoorbeeld geen suberine. Zij worden doorgangscellen genoemd. In sommige plantensoorten ondergaat de endodermis een extra ontwikkelingsfase waarin hun celwanden geligineiseerd worden en de Casparian strepen een U-vorm vertonen. Passagecellen behouden echter dunne celwanden. De voortdurende groei van wortels laat de endodermis afsterven en verdwijnen.
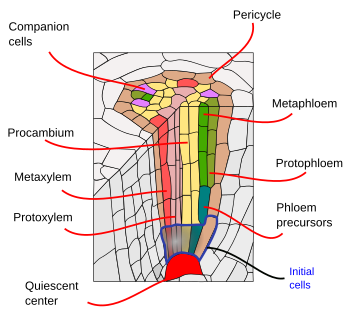
Onder de endodermis bevinden zich een of twee lagen parenchymcellen met zeer dunne celwanden, die de pericyclus vormen. De pericycle cellen kunnen de meristematische activiteit weer op gang brengen en zijwortels vormen. In het oudere deel van de wortels zijn de pericycluscellen sclerefied. In planten met secundaire wortels draagt de pericycle bij tot de vorming van het vasculaire cambium en de phellogen (kurkcambium).
De vaatbundels, xyleem en floëem, bevinden zich in het binnenste deel van de wortel. Primair xyleem en primair floëem zijn in afzonderlijke en afwisselende rijen gerangschikt. Naar gelang van het aantal rijen zijn er diarch (2), triarch (3), tetrarch (4). De tetrarchische organisatie is typisch voor tweezaadlobbigen en gimnospermen (figuur 5). Monocotyledonen zijn polyarctisch met vele rijen xyleem en floëem.

Laterale wortels worden na de embryonaire periode gegenereerd en bepalen de morfologie van het wortelsysteem van de plant. De vorming van zijwortels begint in de pericycle nabij de apicale tip van de wortel, in het gebied waar de cellen gedifferentieerd worden. Dit verschilt van de wijze waarop de takken en de bladeren worden gevormd. Takken en bladeren worden gedifferentieerd vanuit oppervlakkige meristemen, op een exogene manier. De plaats van het beginpunt van een zijwortel wordt beïnvloed door de organisatie van de vaatbundel. Zijwortelprimordia worden gevormd in punten tegenovergesteld aan de xylempolen bij dicotyledonen of aan de floëmpolen bij monocotyledonen. Bij sommige soorten werkt ook de endodermis mee aan de vorming van de zijwortels.
Nadele wortels ontstaan na de kieming uit cellen in de buurt van de vaatbundels, hetzij als een normaal proces of na een inductieproces. Ze kunnen ontstaan in stengels, bladeren en wortels.
Bibliografie
Furuta KM, Hellmann E, Helariutta Y. 2014. Moleculaire controle van celspecificatie en celdifferentiatie tijdens procambiale ontwikkeling. Annual review of plant biology. 65:607-638.
Peret B, De Rybel B, Casimiro I, Benkova E, Swarup R, Laplaze L, Beeckman T, Bennett MJ. 2009. Arabidopsis laterale wortelontwikkeling: een opkomend verhaal. Trend in de plantenwetenschap. 14: 399-408.
Salazar-Henao J, Vélez-Bermúdez IC, Schmidth W. 2016. De regulatie en plasticiteit van wortelhaarpatronen en morfogenese. Development. 143: 1848-1858.