Celbiologie 07: Microtubuli en celdeling
Dit zijn aantekeningen uit college 7 van de cursus Celbiologie van Harvard Extension.
In college 6 werden microtubuli geïntroduceerd, en in dit college wordt hun rol bij de celdeling besproken. Hier is een inleidende video:
Overzicht van de celcyclus
De celcyclus – het proces van celdeling en replicatie – wordt bestuurd door een reeks biochemische schakelaars die het cell cycle control system worden genoemd.
De celcyclus is onderverdeeld in fasen die weer onderverdeeld zijn in fasen – mensen zullen verwijzen naar de “4 fasen” maar dan zijn het er eigenlijk 5, en mensen gebruiken ook andere woorden om die fasen te groeperen, en andere woorden om ze onder te verdelen. Ik heb mijn best gedaan om de relatie tussen deze termen samen te vatten in de volgende tabel. (aangepast/uitgebreid van Wikipedia):
| meest algemene groepering | de vermeende “4 fasen” | subfasen |
|---|---|---|
| non-scheidend | Gat 0 (G0) | |
| interfase | Gat 1 (G1) | G1a R G1b |
| Synthese (S) | ||
| Gap 2 (G2) | ||
| Mitose | Mitose (M) | profase prometafase metafase anafase telofase cytokinese |
De inhoud van elke fase wordt prachtig samengevat in deze uitstekende Wikimedia Commons-afbeelding door Kelvinsong:

De snelst delende menselijke cellen kunnen een celcyclus in ongeveer 24 uur voltooien (G1: 9u, S: 10u, G2: 4u, M: 30 min). Gist kan een cyclus in 30 minuten voltooien, en de snelst delende cellen van Drosophila doen er zelfs maar 8 minuten over.
De belangrijkste regelaars van dit proces zijn de cyclines, die het cycline-afhankelijke kinase of CDK reguleren. Denk eraan dat kinasen eiwitten zijn die andere eiwitten fosforyleren. De fosforylering door CDK van zijn doelwitten maakt het mogelijk dat de mitose doorgaat. Om precies te zijn, de rijpingsbevorderende factor of MPF is een obligaat heterodimeer complex dat bestaat uit cycline B en CDK, dat zijn fosforylerende actie alleen uitvoert als beide eiwitten aanwezig zijn.
Rol van microtubuli
Microtubuli zijn van cruciaal belang gedurende de gehele celcyclus – zij ordenen cellulaire componenten en splitsen ze in tweeën. Hier volgt een serie video’s van de celcyclus waarin de rol van microtubuli wordt belicht:
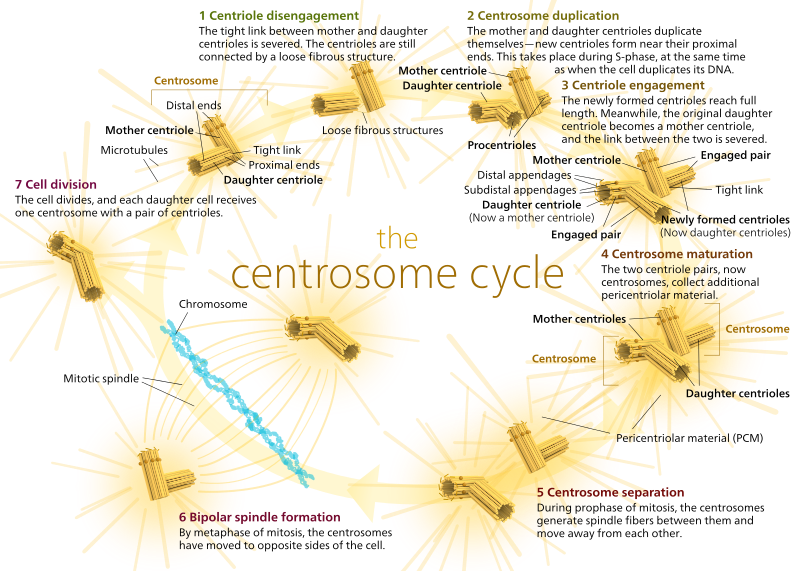
In dieren hebben rustende cellen en zelfs cellen in de interfase meestal maar één centrosoom, dat dient als centraal knooppunt voor alle microtubuli in de cel. Een centrosoom bestaat uit twee centriolen, zoals hieronder is te zien (wederom met dank aan Kelvinsong):

De twee centriolen maken zich los van elkaar en repliceren zich tijdens de S-fase, en scheiden zich vervolgens om tegenovergestelde ‘polen’ van de cel te vormen tijdens de M-fase, zodat er nu twee MTOC’s zijn, die elk uiteindelijk het enige MTOC van een nieuwe cel zullen zijn (nog een afbeelding van de baas Kelvinsong):
Tijdens de mitose heb je dan de twee ‘polen’ van de cel, elk met microtubuli verankerd aan het (-) uiteinde en met hun (+) uiteinden overlappend, wijzend naar het centrum van de cel, zoals hier te zien is (Wikimedia Commons-afbeelding door Lordjuppiter):
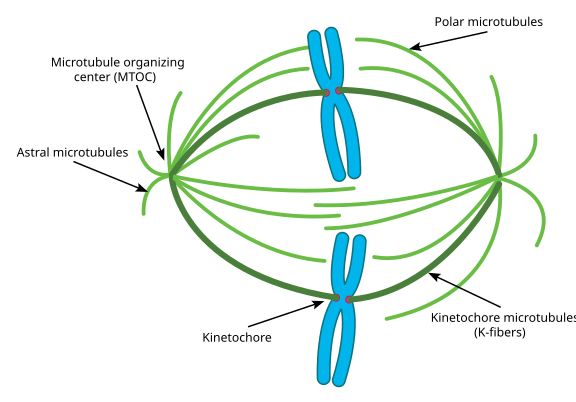
Dat geheel wordt een spindelapparaat genoemd, en het gebied waar de microtubuli van de twee MTOC’s elkaar overlappen wordt de ‘interdigitatiezone’ genoemd. Je hoort soms dat elke MTOC en zijn egelachtige array van microtubuli een ‘mitotische aster’ wordt genoemd.’
Microtubuli in deze fase zouden in drie categorieën vallen:
- Astrale microtubuli wijzen naar buiten, in de richting van de celcortex, om het hele spindelapparaat langs de as van de celdeling te verankeren.
- Kinetochore microtubuli hechten zich aan de kinetochore van chromatiden.
- Polaire microtubuli, evenwijdig aan elkaar maar in tegengestelde richting georiënteerd, zijn van cruciaal belang voor het uit elkaar duwen van het spindelapparaat tijdens de mitose. (In feite zijn polaire microtubuli ook al eerder aanwezig en helpen ze de centrosomen tijdens de profase uit elkaar te duwen).
Als u liever foto’s ziet dan diagrammen, ziet u hier hoe het hele spindelapparaat eruit ziet, met chromatiden in blauw, microtubuli in groen, en de kinetochores als rode stippen:

Microtubuli worden veel dynamischer tijdens mitose: meer gamma-tubuline bevordert gemakkelijker nucleatie, maar XMAP215, een microtubulusstabilisator, wordt tijdens mitose gefosforyleerd en dus geïnactiveerd, waardoor Kinesin-13 vrij blijft om de microtubuli te catastroferen. Fortuinen worden snel gemaakt en verloren. De halfwaardetijd van een microtubule tijdens mitose is ongeveer 15 minuten, vergeleken met 30 minuten tijdens de interfase. Men bestudeert de microtubule-dynamica met behulp van FRAP: men voegt een fluorescerende microtubule toe, laat deze verbleken en kijkt hoe snel de hermontage plaatsvindt aan de hand van hoe snel de fluorescentie weer verschijnt. +Tips spelen ook een belangrijke rol bij het helpen en assembleren van de microtubuli.
Kinesin-5 heeft twee polaire koppen die zich aan tegenover elkaar liggende microtubuli binden en proberen naar het (+) uiteinde van elk te lopen. Dit duwt de twee microtubuli uit elkaar en levert de drijvende kracht voor de scheiding van de MTOCs.
Centromerisch DNA heeft een lage informatie-entropie en speciale histonen die verschillen van ander chromatine. Centromeren zijn een deel van het genoom dat je bijna nooit oppikt bij next-gen sequencing, zelfs niet op zeer grote diepte. Dat komt omdat centromeren een ander doel dienen dan een groot deel van de rest van het genoom: de sequentie daar is gunstig voor interactie met centromereiwitten en kinetochore-hechting. Cohesines zijn eiwitten die de twee zusterchromatiden bij elkaar houden. We zullen kinetochore-eiwitten aanduiden met twee lagen, de binnenste kinetochore en de buitenste kinetochore.
Tijdens de prometafase bewegen de chromosomen heen en weer. Kinesinesines verankeren de chromosomen aan de kinetochore microtubuli voorbij de top, waar Kinesin-13 de microtubuli depolymeriseert, geholpen door een tekort aan beschikbare tubuline-dimeren. Een combinatie van motoreiwitten, op microtubuli inwerkende eiwitten en treadmilling zorgt voor de verplaatsing van de chromosomen. Intussen werken dyneïne en dynactine – motoreiwitten die naar het (-) uiteinde lopen – op de astrale microtubuli en trekken de MTOC’s naar de periferie van de cel. In de metafase komen de chromatiden op één lijn te liggen langs de ‘metafaseplaat’.
Tijdens dit proces lost de nucleaire envelop op, waardoor nucleaire import irrelevant wordt. Ran-GEF lokaliseert zich in de buurt van chromosomen en genereert hoge concentraties Ran-GTP, dat energie levert voor een aantal noodzakelijke processen (?).
Cellen hebben een of ander mechanisme om de spanning in de microtubuli te detecteren die de aanhechting van chromatiden aangeeft voordat de mitose kan doorgaan. Ervoor zorgen dat elk chromatide goed verankerd is, is van cruciaal belang om aneuploïdie te voorkomen.
Naast microtubuli spelen overigens ook andere cytoskeletelementen een sleutelrol in de celcyclus. In de cytokinese vormt actine een samentrekkende ring en knijpt met behulp van myosine II-motoreiwitten de cel in tweeën.
Belang van modelorganismen
De ontdekking van celcyclusregulerende processen berustte sterk op enkele fraaie eigenschappen van populaire modelorganismen.
Saccharomyces cerevisiae (ontluikende gist) en Schizosaccharomyces pombe (splijtgist) kunnen bestaan als haploïden of diploïden. Dat is belangrijk omdat in de haploïde fase één mutatie een gen kan uitschakelen – je hoeft niet beide allelen te raken. En in gist zijn veel mutaties, vooral in de Cdc__ (cell division control) genen, temperatuurafhankelijk, waarbij een eiwit met een missense mutatie nog goed kan functioneren bij ‘permissieve’ temperaturen, maar zijn eigen functie verliest bij ‘niet-permissieve’ temperaturen. Dit maakt het mogelijk het knock-outfenotype te bestuderen (bij de niet-permissieve temperatuur), terwijl de organismen toch gemakkelijk kunnen worden vermeerderd (bij de permissieve temperatuur). Het volledige genoom van S. cerevisiae is beschikbaar in de vorm van plasmidenbibliotheken, zodat kan worden nagegaan welk plasmide het fenotype van een bepaalde mutant herstelt. Zo zijn veel van de genen ontdekt die de celcyclus regelen
In S. cerivisiae maakt budding deel uit van fase G1, en zodra de dochtercel een bepaalde grootte heeft bereikt, op een moment dat “START” wordt genoemd, zijn de twee gecommitteerd om S binnen te gaan en uiteindelijk de celcyclus te voltooien. Zoogdiercellen hebben hun eigen bindingspunt, het restrictiepunt of R, in G1, dat analoog is aan START.
Temperatuurgevoelige Cdc28-mutanten ontluiken niet bij de niet-permissieve temperatuur. Het Cdc28-gen codeert voor gist’s homoloog van ons cycline-afhankelijk kinase (CDK) dat, wanneer en alleen wanneer het een complex vormt met cycline, andere eiwitten kan fosforyleren om hun deelname aan de fasen van de celcyclus te reguleren. Temperatuurgevoelige mutanten blijven bij de niet-permissieve temperatuur steken, niet in staat om te ontluiken en in de S-fase te komen. In plaats daarvan gedragen zij zich als wild-type cellen die van voedingsstoffen verstoken blijven: zij worden groot genoeg om START te passeren, maar gaan dan niet verder.
Xenopus (een soort kikker) is van cruciaal belang gebleken voor het begrijpen van de celcyclus, omdat bij de voortplanting ervan een zeer groot aantal cellen betrokken is (d.w.z. voldoende uitgangsmateriaal voor Western blots, enz.) die perfect gesynchroniseerd zijn (d.w.z. dat zij zich alle op hetzelfde moment in dezelfde fase van de celcyclus bevinden. (Vergelijk dit bijvoorbeeld met gist, waar de cellen niet allemaal op hetzelfde moment in dezelfde fase zijn). Ook het ei zelf is groot en gemakkelijk te bewerken, en meerdere celcycli volgen op de bevruchting. Bij kikkers beginnen de eicellen met de meiotische deling, maar blijven dan 8 maanden in de G2-fase steken, terwijl ze groter worden en een voorraad aanleggen die nodig is voor de groei na de bevruchting.
Tussenliggende filamenten
Naast microfilamenten en microtubuli hebben eukaryote cellen ook een hele reeks ‘andere’ cytoskelet-eiwitten, die tussenliggende filamenten (IF’s) worden genoemd. Hoewel ze diverser zijn dan microfilamenten en microtubuli, zijn IF’s niet zomaar een containerbegrip voor ‘elk ander filament’ – het is eerder een groep verwante eiwitten. Zij strekken zich over het algemeen uit door het cytoplasma en de binnenste kernschil, zijn niet polair en hebben geen met hen geassocieerde motoreiwitten. Zij hebben een grote treksterkte en zijn zeer stabiel, met een langzame uitwisselingssnelheid en niet veel afbraak, hoewel fosforylering hun demontage kan bevorderen. Hier zijn enkele populaire voorbeelden:
- Keratines worden gevonden in epitheelcellen, mesodermcellen en neuronen. Ze geven stevigheid en bestaan in zure en basische vorm. Ze kunnen elk een eigen streng vormen, maar de meeste IF’s bestaan uit twee strengen – een basische en een zure, als het ware om elkaar heen gedraaid. Haar en nagels zijn gemaakt van ‘harde’ keratine, rijk aan cysteïne voor disulfidebindingen, die voor de immense sterkte zorgen. Permanent en steil maken berust op het verminderen van de disulfidebindingen, het hervormen van het haar en vervolgens het hervormen van de disulfidebindingen. Je hebt ook ‘zachte’ keratine in je huid.
- Desmins zoals vimentine worden gevonden in mesenchymale cellen (bot, kraakbeen en vet).
- Neurofilamenten zitten in neuronale axonen en regelen de diameter daarvan, die op zijn beurt de snelheid van de voortplanting van actiepotentialen bepaalt.
- Lamins zijn zowel het meest verbreid als worden verondersteld het meest te lijken op de fylogenetische voorouder van alle andere IF’s. Zij bieden structurele steun voor het kernmembraan. Mogelijk helpen ze de nucleaire poriecomplexen ruimte te geven en organiseren ze ook DNA.
Tot slot een samenvattende video: