Membraanpotentiaal
De membraanpotentiaal in een cel is uiteindelijk het gevolg van twee factoren: elektrische kracht en diffusie. Elektrische kracht ontstaat uit de wederzijdse aantrekking tussen deeltjes met tegengestelde elektrische ladingen (positief en negatief) en de wederzijdse afstoting tussen deeltjes met hetzelfde type lading (beide positief of beide negatief). Diffusie ontstaat door de statistische neiging van deeltjes om zich te herverdelen van gebieden waar ze sterk geconcentreerd zijn naar gebieden waar de concentratie laag is.
SpanningEdit
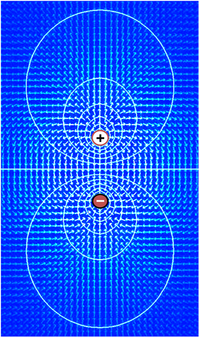
Voltage, dat synoniem is met verschil in elektrische potentiaal, is het vermogen om een elektrische stroom over een weerstand te drijven. De eenvoudigste definitie van een spanning wordt inderdaad gegeven door de wet van Ohm: V=IR, waarbij V spanning is, I stroom en R weerstand. Indien een spanningsbron, zoals een batterij, in een elektrisch circuit wordt geplaatst, zal de hoeveelheid stroom die door de bron over de beschikbare weerstand wordt geleid, groter zijn naarmate de spanning van de bron hoger is. De functionele betekenis van spanning ligt alleen in potentiaalverschillen tussen twee punten in een stroomkring. Het idee van een spanning op een enkel punt is zinloos. Het is in de elektronica gebruikelijk om een spanning van nul toe te kennen aan een willekeurig element van de schakeling, en dan spanningen toe te kennen aan andere elementen gemeten ten opzichte van dat nulpunt. Het is niet van belang welk element als nulpunt wordt gekozen – de functie van een schakeling hangt alleen af van de verschillen, niet van de spanningen op zich. In de meeste gevallen en volgens afspraak wordt het nulpunt echter toegewezen aan het deel van een stroomkring dat in contact staat met de aarde.
Hetzelfde principe geldt voor spanning in de celbiologie. In elektrisch actief weefsel kan het potentiaalverschil tussen twee willekeurige punten worden gemeten door op elk punt een elektrode in te brengen, bijvoorbeeld een binnen en een buiten de cel, en beide elektroden te verbinden met de aansluitdraden van wat in wezen een gespecialiseerde voltmeter is. Bij conventie, wordt de nul potentiaalwaarde toegewezen aan de buitenkant van de cel en het teken van het potentiaalverschil tussen de buitenkant en de binnenkant wordt bepaald door de potentiaal van de binnenkant ten opzichte van de buiten nul.
In wiskundige termen, begint de definitie van spanning met het concept van een elektrisch veld E, een vectorveld dat een grootte en richting toekent aan elk punt in de ruimte. In veel situaties is het elektrisch veld een conservatief veld, hetgeen betekent dat het kan worden uitgedrukt als de gradiënt van een scalaire functie V, d.w.z. E = -∇V. Dit scalaire veld V wordt de spanningsdistributie genoemd. Merk op dat de definitie een willekeurige integratieconstante toelaat – daarom zijn absolute waarden van spanning niet zinvol. In het algemeen kunnen elektrische velden alleen als conservatief worden behandeld als magnetische velden er geen significante invloed op hebben, maar deze voorwaarde geldt meestal goed voor biologisch weefsel.
Omdat het elektrisch veld de gradiënt van de spanningsverdeling is, impliceren snelle veranderingen in spanning binnen een klein gebied een sterk elektrisch veld; omgekeerd, als de spanning over een groot gebied ongeveer gelijk blijft, moeten de elektrische velden in dat gebied zwak zijn. Een sterk elektrisch veld, gelijkwaardig aan een sterke spanningsgradiënt, impliceert dat een sterke kracht wordt uitgeoefend op alle geladen deeltjes die binnen het gebied liggen.
Ionen en de krachten die hun beweging aandrijvenEdit

Elektrische signalen binnen biologische organismen worden in het algemeen aangedreven door ionen. De belangrijkste kationen voor de actiepotentiaal zijn natrium (Na+) en kalium (K+). Dit zijn beide monovalente kationen die één positieve lading hebben. Actiepotentialen kunnen ook calcium (Ca2+) omvatten, een tweewaardig kation dat een dubbele positieve lading heeft. Het chloride-anion (Cl-) speelt een belangrijke rol in de actiepotentialen van sommige algen, maar speelt een te verwaarlozen rol in de actiepotentialen van de meeste dieren.
Ionen passeren het celmembraan onder twee invloeden: diffusie en elektrische velden. Een eenvoudig voorbeeld waarin twee oplossingen-A en B- worden gescheiden door een poreuze barrière illustreert dat diffusie ervoor zal zorgen dat zij zich uiteindelijk zullen vermengen tot gelijke oplossingen. Dit mengen gebeurt door het verschil in hun concentraties. Het gebied met de hoge concentratie zal diffunderen naar het gebied met de lage concentratie. Om het voorbeeld uit te breiden: laat oplossing A 30 natriumionen en 30 chloride-ionen bevatten. En oplossing B heeft maar 20 natriumionen en 20 chloride-ionen. Ervan uitgaande dat de barrière beide soorten ionen doorlaat, dan wordt een stationaire toestand bereikt waarbij beide oplossingen 25 natriumionen en 25 chloride-ionen hebben. Als de poreuze barrière echter selectief is voor wat betreft de ionen die worden doorgelaten, dan zal diffusie alleen niet bepalend zijn voor de resulterende oplossing. Om terug te komen op het vorige voorbeeld, laten we nu een barrière construeren die alleen doorlaatbaar is voor natriumionen. Nu mag alleen natrium door de barrière diffunderen van de hogere concentratie in oplossing A naar de lagere concentratie in oplossing B. Dit zal resulteren in een grotere opeenhoping van natriumionen dan chloride-ionen in oplossing B en een kleiner aantal natriumionen dan chloride-ionen in oplossing A.
Dit betekent dat er een netto positieve lading is in oplossing B door de hogere concentratie van positief geladen natriumionen dan negatief geladen chloride-ionen. Evenzo is er een netto negatieve lading in oplossing A door de grotere concentratie van negatieve chloride-ionen dan positieve natriumionen. Aangezien tegengestelde ladingen elkaar aantrekken en gelijke ladingen elkaar afstoten, worden de ionen nu ook beïnvloed door elektrische velden en diffusiekrachten. Daarom zullen de positieve natriumionen minder geneigd zijn om naar de nu meer positieve B-oplossing te gaan en in de nu meer negatieve A-oplossing te blijven. Het punt waarop de krachten van de elektrische velden de diffusiekracht volledig tegenwerken, wordt de evenwichtspotentiaal genoemd. Op dit punt is de netto stroom van het specifieke ion (in dit geval natrium) nul.
PlasmamembranenEdit

Iedere cel is omgeven door een plasmamembraan, dat de structuur heeft van een lipidenbilaag met daarin vele soorten grote moleculen ingebed. Omdat het uit lipidemoleculen bestaat, heeft het plasmamembraan intrinsiek een hoge elektrische weerstand, met andere woorden een lage intrinsieke permeabiliteit voor ionen. Sommige moleculen in het membraan zijn echter in staat ionen actief van de ene kant van het membraan naar de andere te transporteren of kanalen te vormen waardoor ze kunnen bewegen.
In elektrische terminologie functioneert het plasmamembraan als een gecombineerde weerstand en condensator. De weerstand komt voort uit het feit dat het membraan de beweging van ladingen over het membraan belemmert. Capaciteit ontstaat doordat de lipide bilaag zo dun is dat een opeenhoping van geladen deeltjes aan één kant leidt tot een elektrische kracht die tegengesteld geladen deeltjes naar de andere kant trekt. De capaciteit van het membraan wordt relatief niet beïnvloed door de moleculen die erin zijn ingebed, zodat deze een min of meer onveranderlijke waarde heeft die op ongeveer 2 μF/cm2 wordt geschat (de totale capaciteit van een stukje membraan is evenredig met de oppervlakte ervan). De geleiding van een zuivere lipide bilaag is daarentegen zo laag, dat deze in biologische situaties altijd wordt gedomineerd door de geleiding van alternatieve routes die door ingebedde moleculen worden verschaft. De capaciteit van het membraan ligt dus min of meer vast, maar de weerstand is zeer variabel.
De dikte van een plasmamembraan wordt geschat op ongeveer 7-8 nanometer. Omdat het membraan zo dun is, is er geen zeer grote transmembraanspanning nodig om er een sterk elektrisch veld in op te wekken. Typische membraanpotentialen in dierlijke cellen zijn in de orde van grootte van 100 millivolt (dat is een tiende van een volt), maar berekeningen tonen aan dat dit een elektrisch veld genereert dat dicht bij het maximum ligt dat het membraan kan verdragen – er is berekend dat een spanningsverschil veel groter dan 200 millivolt diëlektrische afbraak zou kunnen veroorzaken, dat is vonkvorming over het membraan.
Gefaciliteerde diffusie en transportEdit

De weerstand van een zuivere lipide bilaag tegen de passage van ionen is zeer hoog, maar structuren die in het membraan zijn ingebed, kunnen de ionenbeweging sterk bevorderen, actief of passief, via mechanismen die transport en diffusie worden vergemakkelijkt. De twee soorten structuren die de grootste rol spelen zijn ionenkanalen en ionenpompen, die beide gewoonlijk bestaan uit assemblages van eiwitmoleculen. Ionenkanalen vormen doorgangen waardoor ionen kunnen bewegen. In de meeste gevallen is een ionkanaal alleen doorlaatbaar voor specifieke soorten ionen (bijvoorbeeld natrium en kalium, maar niet chloride of calcium), en soms varieert de doorlaatbaarheid afhankelijk van de richting van de ionenbeweging. Ionenpompen, ook wel ionentransporteurs of transporteiwitten genoemd, transporteren specifieke soorten ionen actief van de ene kant van het membraan naar de andere, soms met behulp van energie afkomstig van stofwisselingsprocessen om dit te doen.
IonenpompenEdit
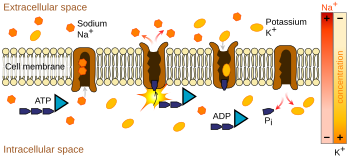
Ionpompen zijn integrale membraaneiwitten die actief transport uitvoeren, d.w.z. cellulaire energie (ATP) gebruiken om de ionen tegen hun concentratiegradiënt in te “pompen”. Dergelijke ionenpompen nemen ionen op aan de ene kant van het membraan (waardoor de concentratie daar daalt) en geven ze af aan de andere kant (waardoor de concentratie daar stijgt).
De ionenpomp die het meest relevant is voor de actiepotentiaal is de natrium-kaliumpomp, die drie natriumionen de cel uit transporteert en twee kaliumionen de cel in. Als gevolg hiervan is de concentratie van kaliumionen K+ in het neuron ruwweg 20-maal zo hoog als de buitenconcentratie, terwijl de natriumconcentratie buiten ruwweg negenmaal zo hoog is als binnen. Op soortgelijke wijze hebben andere ionen verschillende concentraties binnen en buiten het neuron, zoals calcium, chloride en magnesium.
Als de aantallen van elk type ion gelijk zouden zijn, zou de natrium-kaliumpomp elektrisch neutraal zijn, maar door de drie-voor-twee-uitwisseling geeft hij voor elke cyclus een netto verplaatsing van één positieve lading van intracellulair naar extracellulair, waardoor hij bijdraagt tot een positief spanningsverschil. De pomp heeft drie effecten: (1) hij maakt de natriumconcentratie hoog in de extracellulaire ruimte en laag in de intracellulaire ruimte; (2) hij maakt de kaliumconcentratie hoog in de intracellulaire ruimte en laag in de extracellulaire ruimte; (3) hij geeft de intracellulaire ruimte een negatieve spanning ten opzichte van de extracellulaire ruimte.
De natrium-kaliumpomp werkt betrekkelijk langzaam. Als een cel zou worden geïnitialiseerd met overal gelijke concentraties natrium en kalium, zou het uren duren voordat de pomp een evenwicht heeft bereikt. De pomp werkt constant, maar wordt steeds minder efficiënt naarmate de concentraties natrium en kalium die voor pompen beschikbaar zijn, afnemen.
Ionpompen beïnvloeden de actiepotentiaal alleen door de relatieve verhouding tussen intracellulaire en extracellulaire ionenconcentraties vast te stellen. De actiepotentiaal bestaat hoofdzakelijk uit het openen en sluiten van ionenkanalen, niet van ionenpompen. Als de ionenpompen worden uitgeschakeld door hun energiebron weg te nemen, of door een remmer zoals ouabain toe te voegen, kan het axon nog steeds honderdduizenden actiepotentialen afvuren voordat hun amplitudes significant beginnen te dalen. In het bijzonder spelen de ionenpompen geen rol van betekenis bij de repolarisatie van het membraan na een actiepotentiaal.
Een andere functioneel belangrijke ionenpomp is de natrium-calcium wisselaar. Deze pomp werkt op een conceptueel vergelijkbare manier als de natrium-kaliumpomp, behalve dat hij in elke cyclus drie Na+ uit de extracellulaire ruimte uitwisselt tegen één Ca++ uit de intracellulaire ruimte. Omdat de netto stroom van lading inwaarts is, loopt deze pomp in feite “bergafwaarts”, en heeft daarom geen energiebron nodig, behalve de membraanspanning. Het belangrijkste effect van deze pomp is het naar buiten pompen van calcium – het laat ook een inwaartse stroom van natrium toe, waardoor de natrium-kaliumpomp wordt tegengewerkt, maar omdat de totale natrium- en kaliumconcentraties veel hoger zijn dan de calciumconcentraties, is dit effect relatief onbelangrijk. Het netto resultaat van de natrium-calcium wisselaar is dat in de rusttoestand de intracellulaire calciumconcentraties zeer laag worden.
IonenkanalenEdit

Ionkanalen zijn integrale membraaneiwitten met een porie waardoor ionen zich kunnen verplaatsen tussen de extracellulaire ruimte en het celinterieur. De meeste kanalen zijn specifiek (selectief) voor één ion; zo worden de meeste kaliumkanalen gekenmerkt door een selectiviteitsverhouding van 1000:1 voor kalium boven natrium, hoewel kalium- en natriumionen dezelfde lading hebben en slechts in geringe mate in straal van elkaar verschillen. De kanaalporie is doorgaans zo klein dat ionen er in een enkele volgorde doorheen moeten. Kanaalporiën kunnen open of gesloten zijn voor ionpassage, hoewel een aantal kanalen verschillende sub-geleidingsniveaus vertonen. Wanneer een kanaal open is, permeëren ionen door de kanaalporie langs de transmembraan concentratiegradiënt voor dat specifieke ion. De snelheid van de ionenstroom door het kanaal, d.w.z. de stroomamplitude van één kanaal, wordt bepaald door de maximale kanaalgeleiding en de elektrochemische drijvende kracht voor dat ion, namelijk het verschil tussen de momentane waarde van de membraanpotentiaal en de waarde van de omkeerpotentiaal.

Een kanaal kan verschillende toestanden hebben (die overeenkomen met verschillende conformaties van het eiwit), maar elk van deze toestanden is ofwel open ofwel gesloten. In het algemeen komt een gesloten toestand overeen met hetzij een samentrekking van de porie, waardoor deze onbegaanbaar wordt voor het ion, hetzij een afzonderlijk deel van het eiwit, waardoor de porie wordt afgesloten. Bijvoorbeeld, het spanningsafhankelijk natriumkanaal ondergaat inactivatie, waarbij een deel van het eiwit in de porie zwaait, waardoor deze wordt afgesloten. Deze inactivering schakelt de natriumstroom uit en speelt een cruciale rol in de actiepotentiaal.
Ion-kanalen kunnen worden ingedeeld naar de manier waarop zij op hun omgeving reageren. De ionenkanalen die betrokken zijn bij de actiepotentiaal zijn bijvoorbeeld spanningsgevoelige kanalen; zij openen en sluiten zich in reactie op de spanning over het membraan. Ligand-geactiveerde kanalen vormen een andere belangrijke klasse; deze ionenkanalen openen en sluiten zich als reactie op de binding van een ligandmolecule, zoals een neurotransmitter. Andere ionenkanalen gaan open en dicht door mechanische krachten. Weer andere ionenkanalen – zoals die van zintuiglijke neuronen – gaan open en dicht als reactie op andere stimuli, zoals licht, temperatuur of druk.
LekkanalenEdit
Lekkanalen zijn het eenvoudigste type ionenkanaal, in die zin dat hun permeabiliteit min of meer constant is. De typen lekkende kanalen die in neuronen het meest van belang zijn, zijn kalium- en chloorkanalen. Zelfs deze zijn niet volkomen constant in hun eigenschappen: Ten eerste zijn de meeste ervan spanningsafhankelijk in die zin dat zij in de ene richting beter geleiden dan in de andere (met andere woorden, het zijn gelijkrichters); ten tweede kunnen sommige ervan worden uitgeschakeld door chemische liganden, hoewel zij geen liganden nodig hebben om te werken.
Ligand-gated channelsEdit
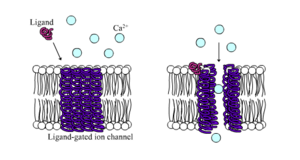
Ligand-gated ionkanalen zijn kanalen waarvan de doorlaatbaarheid sterk wordt verhoogd wanneer een of ander chemisch ligand zich bindt aan de eiwitstructuur. Dierlijke cellen bevatten honderden, zo niet duizenden, typen van deze kanalen. Een groot aantal functioneert als neurotransmitterreceptoren – zij komen voor op postsynaptische plaatsen, en het chemische ligand dat hen activeert wordt afgegeven door het presynaptische axonuiteinde. Een voorbeeld van dit type is de AMPA-receptor, een receptor voor de neurotransmitter glutamaat die, wanneer geactiveerd, natrium- en kaliumionen doorlaat. Een ander voorbeeld is de GABAA-receptor, een receptor voor de neurotransmitter GABA die bij activering chloride-ionen doorlaat.
Neurotransmitterreceptoren worden geactiveerd door liganden die in het extracellulaire gebied voorkomen, maar er zijn andere typen ligand-gated kanalen die door interacties aan de intracellulaire kant worden gestuurd.
Spanningsafhankelijke kanalenEdit
Spanningsafhankelijke ionkanalen, ook wel spanningsafhankelijke ionkanalen genoemd, zijn kanalen waarvan de permeabiliteit wordt beïnvloed door de membraanpotentiaal. Zij vormen een andere zeer grote groep, waarbij elk lid een bepaalde ionenselectiviteit en een bepaalde spanningsafhankelijkheid heeft. Vele zijn ook tijdsafhankelijk, d.w.z. dat zij niet onmiddellijk op een spanningsverandering reageren, maar pas na enige tijd.
Een van de belangrijkste leden van deze groep is een type spanningsafhankelijk natriumkanaal dat aan de basis ligt van actiepotentialen – deze worden ook wel Hodgkin-Huxley natriumkanalen genoemd, omdat zij oorspronkelijk werden gekarakteriseerd door Alan Lloyd Hodgkin en Andrew Huxley in hun Nobelprijswinnende studies naar de fysiologie van de actiepotentiaal. Het kanaal is gesloten op het spanningsniveau in rust, maar opent abrupt wanneer de spanning een bepaalde drempel overschrijdt, waardoor een grote instroom van natriumionen mogelijk wordt die een zeer snelle verandering van de membraanpotentiaal veroorzaakt. Het herstel van een actiepotentiaal is gedeeltelijk afhankelijk van een type spanningsgestuurd kaliumkanaal dat gesloten is bij het rustspanningsniveau, maar zich opent als gevolg van de grote spanningsverandering die tijdens de actiepotentiaal wordt veroorzaakt.
OmkeerpotentiaalEdit
De omkeerpotentiaal (of evenwichtspotentiaal) van een ion is de waarde van de transmembraanspanning waarbij diffusieve en elektrische krachten elkaar compenseren, zodat er geen netto ionenstroom over het membraan is. Dit betekent dat de transmembraanspanning precies tegengesteld is aan de diffusiekracht van het ion, zodat de netto stroom van het ion over het membraan nul en onveranderlijk is. De omkeerpotentiaal is belangrijk omdat het de spanning geeft die werkt op kanalen die doorlaatbaar zijn voor dat ion – met andere woorden, het geeft de spanning die de ionconcentratiegradiënt genereert wanneer deze werkt als een batterij.
De evenwichtspotentiaal van een bepaald ion wordt gewoonlijk aangeduid met de notatie Eion.De evenwichtspotentiaal voor elk ion kan worden berekend met behulp van de Nernst-vergelijking. Bijvoorbeeld, de omkeerpotentiaal voor kaliumionen zal als volgt zijn:
E e q , K + = R T z F ln o i , {Displaystyle E_{eq,K^{+}}={\frac {RT}{zF}}}ln {\frac {_{o}}{_{i}}},}

waar
- Eeq,K+ is de evenwichtspotentiaal voor kalium, gemeten in volt
- R is de universele gasconstante, gelijk aan 8,314 joule-K-1-mol-1
- T is de absolute temperatuur, gemeten in kelvin (= K = graden Celsius + 273.15)
- z is het aantal elementaire ladingen van het ion in kwestie dat bij de reactie betrokken is
- F is de constante van Faraday, gelijk aan 96,485 coulombs-mol-1 of J-V-1-mol-1
- o is de extracellulaire concentratie van kalium, gemeten in mol-m-3 of mmol-l-1
- i is de intracellulaire kaliumconcentratie
Zelfs als twee verschillende ionen dezelfde lading hebben (d.e., K+ en Na+), kunnen zij toch zeer verschillende evenwichtspotentialen hebben, mits hun buiten- en/of binnenconcentraties verschillen. Neem bijvoorbeeld de evenwichtspotentialen van kalium en natrium in neuronen. De kalium-evenwichtspotentiaal EK is -84 mV met 5 mM kalium buiten en 140 mM binnen. Aan de andere kant is de natrium evenwichtspotentiaal, ENa, ongeveer +66 mV met ongeveer 12 mM natrium binnen en 140 mM buiten.
Veranderingen in membraanpotentiaal tijdens ontwikkelingEdit
De rustmembraanpotentiaal van een neuron verandert in feite tijdens de ontwikkeling van een organisme. Om ervoor te zorgen dat een neuron uiteindelijk zijn volledige volwassen functie kan vervullen, moet zijn potentiaal tijdens de ontwikkeling strak worden gereguleerd. Naarmate een organisme zich verder ontwikkelt, wordt het rustmembraanpotentiaal negatiever. Naarmate de ontwikkeling in de hersenen vordert, differentiëren en vermenigvuldigen zich ook gliacellen. De toevoeging van deze gliacellen verhoogt het vermogen van het organisme om het extracellulaire kalium te reguleren. De daling van extracellulair kalium kan leiden tot een daling van de membraanpotentiaal met 35 mV.
Cell excitabilityEdit
Cel exciteerbaarheid is de verandering in membraanpotentiaal die noodzakelijk is voor cellulaire reacties in verschillende weefsels. Exciteerbaarheid van cellen is een eigenschap die wordt geïnduceerd tijdens de vroege embriogenese. De exciteerbaarheid van een cel is ook gedefinieerd als het gemak waarmee een reactie kan worden uitgelokt. De rust- en drempelpotentialen vormen de basis van de exciteerbaarheid van cellen en deze processen zijn fundamenteel voor de generatie van gradatie- en actiepotentialen.
De belangrijkste regulatoren van de exciteerbaarheid van cellen zijn de extracellulaire elektrolytconcentraties (d.w.z. Na+, K+, Ca2+, Cl-, Mg2+) en daarmee geassocieerde eiwitten. Belangrijke eiwitten die de exciteerbaarheid van cellen reguleren zijn spanningsafhankelijke ionenkanalen, ionentransporters (b.v. Na+/K+-ATPase, magnesiumtransporters, zuur-base transporters), membraanreceptoren en hyperpolarisatie-geactiveerde cyclisch-nucleotide-geactiveerde kanalen. Zo zijn bijvoorbeeld kaliumkanalen en calciumgevoelige receptoren belangrijke regulatoren van de prikkelbaarheid in neuronen, hartspiercellen en vele andere prikkelgevoelige cellen zoals astrocyten. Calciumion is ook de belangrijkste tweede boodschapper in exciteerbare celsignalering. Activering van synaptische receptoren leidt tot langdurige veranderingen in de neuronale prikkelbaarheid. Schildklier-, bijnier- en andere hormonen reguleren ook de exciteerbaarheid van cellen.
Veel celtypen worden geacht een exciteerbaar membraan te hebben. Prikkelbare cellen zijn neuronen, myocyten (hartcellen, skeletcellen, gladde cellen), vasculaire endotheelcellen, juxtaglomerulaire cellen, interstitiële cellen van Cajal, vele soorten epitheelcellen (b.v. bètacellen, alfacellen, deltacellen, entero-endocriene cellen), gliacellen (bv. astrocyten), mechanoreceptorcellen (bv. haarcellen en Merkelcellen), chemoreceptorcellen (bv. glomuscellen, smaakreceptoren), sommige plantencellen en mogelijk immuuncellen. Astrocyten vertonen een vorm van niet-elektrische exciteerbaarheid op basis van intracellulaire calciumschommelingen die verband houden met de expressie van verschillende receptoren waarmee zij het synaptische signaal kunnen detecteren. In neuronen zijn er verschillende membraaneigenschappen in sommige delen van de cel, bijvoorbeeld, dendritische exciteerbaarheid verleent neuronen het vermogen tot toevallige detectie van ruimtelijk gescheiden inputs.
Equivalente schakelingEdit
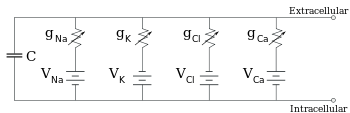
Electrofysiologen modelleren de effecten van ionische concentratieverschillen, ionenkanalen en membraancapaciteit in termen van een equivalent circuit, dat bedoeld is om de elektrische eigenschappen van een kleine patch van het membraan weer te geven. De equivalente stroomkring bestaat uit een condensator parallel met vier banen die elk bestaan uit een batterij in serie met een variabele geleiding. De capaciteit wordt bepaald door de eigenschappen van de lipidenbilaag, en wordt als vast beschouwd. Elk van de vier parallelle banen is afkomstig van een van de belangrijkste ionen, natrium, kalium, chloride, en calcium. De spanning van elke ionenbaan wordt bepaald door de concentraties van het ion aan elke kant van het membraan; zie het gedeelte Omkeerpotentiaal hierboven. De geleiding van elke ionenbaan op een bepaald moment wordt bepaald door de toestand van alle ionenkanalen die potentieel doorlaatbaar zijn voor dat ion, met inbegrip van lekkende kanalen, ligand-geactiveerde kanalen en spanningsgeactiveerde ionenkanalen.
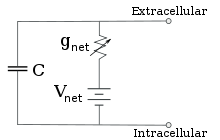
Voor vaste ionconcentraties en vaste waarden van de ionkanaalgeleiding, kan de equivalente stroomkring met behulp van de Goldman-vergelijking, zoals hieronder beschreven, verder worden gereduceerd tot een stroomkring die een capaciteit parallel met een batterij en een geleiding bevat. In elektrische termen is dit een soort RC-kring (weerstand-capaciteit-kring), en de elektrische eigenschappen ervan zijn zeer eenvoudig. Uitgaande van een willekeurige begintoestand neemt de stroom over hetzij de geleiding hetzij de capaciteit exponentieel af, met een tijdconstante van τ = RC, waarbij C de capaciteit van het membraangedeelte is, en R = 1/gnet de nettoweerstand. Voor realistische situaties ligt de tijdconstante meestal in het bereik van 1-100 milliseconden. In de meeste gevallen treden veranderingen in de geleiding van ionenkanalen op een snellere tijdschaal op, zodat een RC-circuit geen goede benadering is; de differentiaalvergelijking die wordt gebruikt om een membraanflard te modelleren is echter meestal een aangepaste versie van de RC-circuitvergelijking.