Atlas histologii roślin i zwierząt
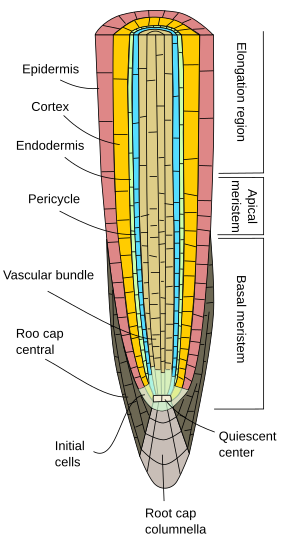
Początkowo korzenie rosną na długość dzięki aktywności merystemu wierzchołkowego korzenia, który jest chroniony przez czapeczkę korzeniową. Proliferacja i różnicowanie komórek merystemu wierzchołkowego korzenia daje komórki, które najpierw są zorganizowane jako tzw. korzeń pierwotny, czyli wykazujący wzrost pierwotny. Jest to organizacja mniej złożona niż łodyga, ponieważ brak w niej węzłów, międzywęźli i liści.
Niezależnie od gatunku rośliny, korzenie pierwotne wykazują epidermę, czyli ryzodermę, zwykle jednozarodnikową (o grubej warstwie komórek), pozbawioną aparatów szparkowych i wykazującą włoski korzeniowe w regionie dojrzewania, zaraz po regionie wydłużania (ryc. 1, 2 i 3). Ogólnie rzecz biorąc, w pobliżu wierzchołka korzenia komórki epidermy wykazują cienką kutykulę ułatwiającą wnikanie wody i soli mineralnych. U niektórych gatunków, np. roślin kserofitów, oraz u korzeni położonych blisko powierzchni gleby, pod epidermą znajduje się warstwa komórek zwana hipodermą. Komórki hipodermy tworzą cienką warstwę komórek z suberalizowanymi ścianami komórkowymi. Hipoderma może być dalej wyspecjalizowana i staje się egzodermą, która występuje u niektórych roślin okrytozalążkowych i działa jako druga bariera zapobiegająca swobodnej dyfuzji substancji między korzeniem a glebą.

Włośniki korzeniowe absorbują wodę i sole mineralne. Są to wydłużone komórki epidermalne występujące w regionie dojrzewania, które zwiększają powierzchnię absorpcyjną w kontakcie ze środowiskiem zewnętrznym, a tym samym zdolność absorpcyjną. Włoski korzeniowe tworzą się i zanikają w sposób ciągły w miarę wzrostu korzenia, ponieważ region dojrzewania podąża za rosnącym wierzchołkiem korzenia w mniej więcej stałej odległości. Liczba włośników korzeniowych wynosi od 20 do 500 na cm2 w korzeniach drzew i około 25000 na cm2 w korzeniach żyta ozimego. Liczba ta może się także zmieniać w zależności od warunków środowiska.
Wyróżnia się trzy wzorce organizacji włośników (rys. 4). Gęstość włośników zależy od warunków środowiska. Na przykład, w glebach o niskiej zawartości fosforanów liczba włośników jest większa, aby zwiększyć całkowitą powierzchnię absorpcji korzeni. Wiele organizmów symbiotycznych, takich jak bakterie wiążące azot, jest związanych z włośnikami.


Kora korzenia znajduje się poniżej epidermy, czyli hipodermy. W korzeniach, kora jest gruba lub bardzo gruba (znacznie bardziej niż w łodygach), i jest zazwyczaj zbudowana z komórek parenchymy wyspecjalizowanych w magazynowaniu, chociaż może to być parenchyma fotosyntetyczna w korzeniach powietrznych i parenchyma aerofitowa w korzeniach wodnych. Pomiędzy komórkami parenchymy korowej znajduje się wiele pustych przestrzeni, dzięki którym woda może być odprowadzana (droga apoplastyczna) w kierunku wiązki naczyniowej. Ponadto, komórki parenchymy są połączone między sobą plazmodesmami (droga symplastowa), które umożliwiają minerałom i wodzie przemieszczanie się z komórki do komórki.

Odrębną cechą wzrostu pierwotnego korzenia jest endoderma, najbardziej wewnętrzna warstwa kory. Endoderma jest ewolucyjnie zachowana od paproci do okrytozalążkowych. Jest to jednokomórkowa warstwa ściśle zbitych komórek, których ściany komórkowe są częściowo impregnowane suberyną i tworzą zgrubienia, tzw. paski Caspariego. Zagęszczenie komórek i nieprzepuszczalność pasków Caspariego sprawia, że woda i substancje rozpuszczone nie mogą przechodzić przez endodermę przez cytoplazmę komórek endodermy. W ten sposób endoderma stanowi barierę przed wolną dyfuzją, która kontroluje przemieszczanie się substancji z podłoża do wiązek naczyniowych. W tych regionach korzenia, w których następuje wzrost wtórny, zarówno endoderma, jak i kora ulegają zanikowi (patrz następna strona).
Paski Kaspariana to impregnaty ściany pierwotnej, które otaczają komórki endodermy jako pas ułożony podłużnie. Nie są one ścianą wtórną. Paski są ciągłe pomiędzy sąsiednimi komórkami poprzez blaszkę środkową. W widoku 3D przypominają cylindryczną sieć rybacką, w której sznurki to paski Caspariana, a otwory to komórki endodermy. Paski Caspariana zawierają ligninę, ale nie suberynę. Suberyna może być jednak obecna w ścianie komórkowej komórek endodermy po uformowaniu się pasków Caspariego. Suberyna jest ułożona w postaci warstw na powierzchni ściany komórkowej, o różnej grubości w zależności od lokalizacji komórki. Na przykład komórki endodermy znajdujące się bliżej biegunów ksylemu nie syntetyzują suberyny. Nazywa się je komórkami przejściowymi. U niektórych gatunków roślin endoderma przechodzi dodatkowy etap rozwoju, w którym ich ściany komórkowe ulegają zdrewnieniu, a paski Caspariana przybierają kształt litery U. Komórki pasażu zachowują jednak cienkie ściany komórkowe. Ciągły wzrost korzeni prowadzi do obumierania i zaniku endodermy.
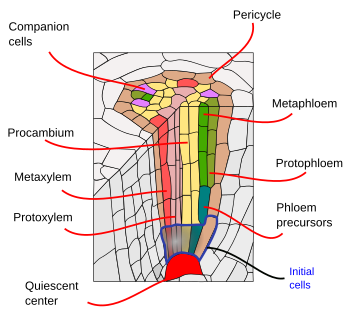
Poniżej endodermy znajduje się jedna lub dwie warstwy komórek parenchymy o bardzo cienkich ścianach komórkowych, które tworzą perycykl. Komórki perycyklu mogą wznowić aktywność merystematyczną i uformować korzenie boczne. W starszej części korzenia komórki perycyklu są sklerotyzowane. U roślin z korzeniem wtórnym perycykl bierze udział w tworzeniu kambium naczyniowego i phellogenu (kambium korkowego).
Wiązki naczyniowe, ksylem i łyko, znajdują się w wewnętrznej części korzenia. Ksylem pierwotny i łyko pierwotne ułożone są w oddzielnych, naprzemiennych rzędach. Ze względu na liczbę rzędów wyróżnia się diarchię (2), triarchię (3), tetrarchię (4). Organizacja tetrarchiczna jest typowa dla dwuliściennych i gimnospermy (rys. 5). Jednoliścienne są wieloskośne, mają wiele rzędów ksylemu i łyka.

Korzenie boczne powstają po okresie embrionalnym i decydują o morfologii systemu korzeniowego rośliny. Tworzenie korzeni bocznych rozpoczyna się w perycyklu w pobliżu wierzchołka korzenia, w regionie, w którym komórki stają się zróżnicowane. Różni się to od sposobu powstawania gałęzi i liści. Gałęzie i liście różnicują się z merystemów powierzchniowych, w sposób egzogeniczny. Na położenie punktu startowego korzenia bocznego ma wpływ organizacja wiązki naczyniowej. Pierwotniaki korzeni bocznych powstają w miejscach przeciwległych do biegunów ksylemu u dwuliściennych lub do biegunów łyka u jednoliściennych. U niektórych gatunków endoderma współdziała również w tworzeniu korzeni bocznych.
Korzenie przybyszowe powstają po wykiełkowaniu z komórek w pobliżu wiązek naczyniowych, jako proces normalny lub po indukcji. Mogą powstawać w łodygach, liściach i korzeniach.
Bibliografia
Furuta KM, Hellmann E, Helariutta Y. 2014. Molekularna kontrola specyfikacji komórek i różnicowania komórkowego podczas rozwoju prokambialnego. Annual review of plant biology. 65:607-638.
Peret B, De Rybel B, Casimiro I, Benkova E, Swarup R, Laplaze L, Beeckman T, Bennett MJ. 2009. Arabidopsis lateral root development: an emerging story. Trend in plant science. 14: 399-408.
Salazar-Henao J, Vélez-Bermúdez IC, Schmidth W. 2016. The regulation and plasticity of root hair patterning and morphogenesis. Development. 143: 1848-1858.