Biologia komórki w ekspresji molekularnej: Filamenty pośrednie
Filamenty pośrednie
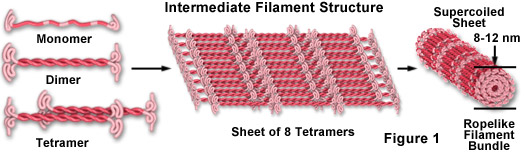
Filamenty pośrednie to bardzo szeroka klasa białek włóknistych, które odgrywają ważną rolę zarówno jako strukturalne, jak i funkcjonalne elementy cytoszkieletu. W zakresie wielkości od 8 do 12 nanometrów (w średnicy; patrz Rycina 1), filamenty pośrednie funkcjonują jako elementy przenoszące napięcie, pomagając w utrzymaniu kształtu i sztywności komórki, a także służą do zakotwiczenia w miejscu kilku organelli, w tym jądra i desmosomów. Filamenty pośrednie biorą również udział w tworzeniu blaszki jądrowej, siatkowatego układu, który pokrywa wewnętrzną błonę jądrową i reguluje kształt jądra.
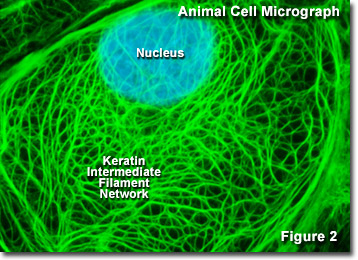
Ale wszystkie eukarionty zawierają wspólne elementy cytoszkieletowe aktynę i tubulinę (zarówno wolne w cytoplazmie, jak i spolimeryzowane w postaci mikrofilamentów i mikrotubul), filamenty pośrednie występują tylko u niektórych gatunków metazoan, m.in. u kręgowców, nicieni i mięczaków. U kręgowców obecność i skład filamentów pośrednich jest nie tylko zależna od gatunku, ale także zmienia się w zależności od typu tkanki. Na przykład, większość komórek nabłonkowych zwierząt zawiera keratyny, zróżnicowaną rodzinę filamentów pośrednich składającą się z ponad 50 członków, podczas gdy komórki mezenchymalne i mięśniowe są bogate w białka włókniste, odpowiednio wimentyny i desminy. Filamenty pośrednie występujące w neuronach i komórkach glejowych obejmują peryferynę, neurofilamenty i białko kwasowe fibryli glejowych (GFAP). Różnorodne białka towarzyszące wiążą się z filamentami pośrednimi w celu poprawy stabilności (poprzez sieciowanie) lub zapewnienia miejsc przyczepu dla innych zespołów białek, takich jak filamenty aktynowe i mikrotubule.
Inną wysoce wyspecjalizowaną klasą filamentów pośrednich są laminy jądrowe, które tworzą włóknistą sieć białkową, która pokrywa wnętrze błony jądrowej, jak omówiono powyżej. Ze względu na ich bliskie sąsiedztwo z błoną, laminy jądrowe pomagają przytwierdzić chromosomy do błony jądrowej i zapewniają punkty zakotwiczenia dla porów jądrowych. Wielu naukowców uważa, że laminy jądrowe są ewolucyjnym przodkiem cytoplazmatycznych filamentów pośrednich, które powstały w wyniku duplikacji i translokacji produktu genu do cytoplazmy. Sztywność nadana komórkom przez sieci filamentów pośrednich jest szczególnie przydatna dla zwierząt o miękkim ciele, które nie posiadają egzoszkieletu. Ponieważ filamenty pośrednie występują bardzo obficie w komórkach, które są często poddawane dużym naprężeniom mechanicznym in vivo, wydaje się, że ich podstawową rolą jest zapewnienie fizycznej wytrzymałości komórkom i tkankom.
Jak pokazano na rycinie 1, monomery peptydów filamentu pośredniego są wydłużoną, włóknistą klasą białek z centralnym regionem alfa-helikalnym, zakończonym globularnymi końcami zarówno na końcach aminowych, jak i karboksylowych. Dwie z jednostek monomeru tworzą dimer w kształcie cewki, który samoistnie łączy się w układzie antyrównoległym, tworząc tetramer, który jest analogiczną rozpuszczalną podjednostką dla globularnego monomeru aktyny i heterodimeru tubuliny (występującego swobodnie w cytoplazmie). Jednostki tetrameru łączą się poprzecznie, tworząc arkusz ośmiu równoległych protofilamentów, które są zwinięte w ciasną wiązkę. W każdym ciasno zwiniętym przekroju poprzecznym filamentu pośredniego widoczne są 32 pojedyncze alfa-helikalne peptydy, co sprawia, że filament łatwo się zgina, ale dość trudno go złamać, co tłumaczy wyjątkową sztywność struktury. Chociaż mniej wiadomo o mechanizmie montażu i demontażu filamentów pośrednich, jasne jest, że niektóre klasy są bardzo dynamicznymi strukturami o znacznym tempie obrotu w wielu typach komórek.
Mutacje w genach filamentów pośrednich prowadzą do wielu raczej rzadkich chorób. Na przykład, wadliwe keratyny w tkance skórnej prowadzą do zaburzenia znanego jako epidermolysis bullosa simplex, objawiającego się pęcherzami skórnymi wytwarzanymi nawet przy niewielkim obciążeniu mechanicznym. Podobne choroby pęcherzowe wynikające z mutacji keratyn w innych tkankach dotyczą przełyku, oczu i jamy ustnej. Kilka chorób neurodegeneracyjnych, takich jak stwardnienie zanikowe boczne (ALS lub choroba Lou Gehriga), wiąże się z nieprawidłowościami w sieci filamentów pośrednich (neurofilamentów), a defekty filamentów pośrednich desminy powodują zaburzenia mięśniowe.
POWRÓT DO STRUKTURY KOMÓREK ZWIERZĄT
POWRÓT DO STRUKTURY KOMÓREK ROŚLIN
Pytania lub komentarze? Wyślij nam wiadomość e-mail.
© 1995-2021 by Michael W. Davidson and The Florida State University. Wszystkie prawa zastrzeżone. Żadne obrazy, grafiki, oprogramowanie, skrypty ani aplety nie mogą być powielane ani wykorzystywane w żaden sposób bez zgody właścicieli praw autorskich. Korzystanie z tej strony oznacza zgodę na wszystkie warunki prawne określone przez właścicieli.
Ta strona jest utrzymywana przez nasz
Graphics & Web Programming Team
we współpracy z Optical Microscopy at the
National High Magnetic Field Laboratory.
Ostatnia modyfikacja: Friday, Nov 13, 2015 at 02:18 PM
Access Count Since December 14, 2004: 173927
Mikroskopy dostarczone przez:
![]()
![]()