Potencial de membrana
O potencial de membrana numa célula deriva em última análise de dois factores: força eléctrica e difusão. A força eléctrica resulta da atracção mútua entre partículas com cargas eléctricas opostas (positivas e negativas) e da repulsão mútua entre partículas com o mesmo tipo de carga (ambas positivas ou ambas negativas). A difusão surge da tendência estatística das partículas para redistribuir de regiões onde são altamente concentradas para regiões onde a concentração é baixa.
VoltageEdit
Voltagem, que é sinónimo de diferença de potencial eléctrico, é a capacidade de conduzir uma corrente eléctrica através de uma resistência. De facto, a definição mais simples de uma tensão é dada pela lei de Ohm: V=IR, onde V é tensão, I é corrente e R é resistência. Se uma fonte de tensão tal como uma bateria for colocada num circuito eléctrico, quanto maior for a tensão da fonte, maior será a quantidade de corrente que esta irá conduzir através da resistência disponível. O significado funcional da tensão reside apenas nas diferenças potenciais entre dois pontos de um circuito. A ideia de uma voltagem num único ponto não tem qualquer significado. É convencional em electrónica atribuir uma tensão de zero a algum elemento arbitrariamente escolhido do circuito, e depois atribuir tensões a outros elementos medidos em relação a esse ponto zero. Não há significado em que elemento é escolhido como ponto zero – a função de um circuito depende apenas das diferenças e não das tensões per se. Contudo, na maioria dos casos e por convenção, o nível zero é mais frequentemente atribuído à parte de um circuito que está em contacto com a terra.
O mesmo princípio aplica-se à tensão na biologia celular. No tecido electricamente activo, a diferença potencial entre quaisquer dois pontos pode ser medida inserindo um eléctrodo em cada ponto, por exemplo um dentro e um fora da célula, e ligando ambos os eléctrodos aos condutores do que é, na sua essência, um voltímetro especializado. Por convenção, o valor do potencial zero é atribuído ao exterior da célula e o sinal da diferença de potencial entre o exterior e o interior é determinado pelo potencial do interior em relação ao zero exterior.
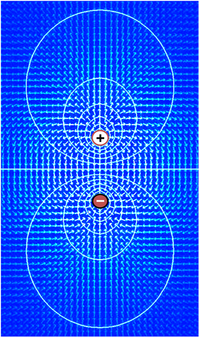
Em termos matemáticos, a definição de tensão começa com o conceito de um campo eléctrico E, um campo vectorial que atribui uma magnitude e direcção a cada ponto no espaço. Em muitas situações, o campo eléctrico é um campo conservador, o que significa que ele pode ser expresso como o gradiente de uma função escalar V, ou seja, E = -∇V. Este campo escalar V é referido como a distribuição de voltagem. Note-se que a definição permite uma constante arbitrária de integração – é por isso que os valores absolutos de voltagem não são significativos. Em geral, os campos eléctricos só podem ser tratados como conservadores se os campos magnéticos não os influenciarem significativamente, mas esta condição aplica-se geralmente bem ao tecido biológico.
Porque o campo eléctrico é o gradiente da distribuição de voltagem, mudanças rápidas de voltagem dentro de uma pequena região implicam um campo eléctrico forte; ao contrário, se a voltagem se mantiver aproximadamente a mesma numa grande região, os campos eléctricos nessa região devem ser fracos. Um campo eléctrico forte, equivalente a um forte gradiente de voltagem, implica que uma força forte é exercida sobre quaisquer partículas carregadas que se encontrem dentro da região.
Iões e as forças que impulsionam o seu movimentoEditar

Os sinais eléctricos dentro dos organismos biológicos são, em geral, impulsionados por iões. Os catiões mais importantes para o potencial de acção são sódio (Na+) e potássio (K+). Ambos são catiões monovalentes que transportam uma única carga positiva. Os potenciais de acção também podem envolver cálcio (Ca2+), que é um catião divalente que carrega uma carga positiva dupla. O anião cloreto (Cl-) desempenha um papel importante nos potenciais de acção de algumas algas, mas desempenha um papel insignificante nos potenciais de acção da maioria dos animais.
Iões atravessam a membrana celular sob duas influências: difusão e campos eléctricos. Um exemplo simples em que duas soluções-A e B estão separadas por uma barreira porosa ilustra que a difusão assegurará que acabarão por se misturar em soluções iguais. Esta mistura ocorre devido à diferença nas suas concentrações. A região com alta concentração irá difundir-se para a região com baixa concentração. Para ampliar o exemplo, deixar a solução A ter 30 iões de sódio e 30 iões de cloreto. Além disso, deixar a solução B ter apenas 20 iões de sódio e 20 iões de cloreto. Assumindo que a barreira permite que ambos os tipos de iões viajem através dela, então será alcançado um estado estável em que ambas as soluções tenham 25 iões de sódio e 25 iões de cloreto. Se, no entanto, a barreira porosa for selectiva para a qual os iões são deixados passar, então a difusão por si só não irá determinar a solução resultante. Voltando ao exemplo anterior, vamos agora construir uma barreira que é permeável apenas aos iões de sódio. Agora, apenas o sódio é permitido difundir através da barreira desde a sua maior concentração na solução A até à menor concentração na solução B. Isto resultará numa maior acumulação de iões sódio do que iões cloreto na solução B e num menor número de iões sódio do que iões cloreto na solução A.
Isto significa que existe uma carga líquida positiva na solução B a partir da maior concentração de iões sódio com carga positiva do que de iões cloreto com carga negativa. Da mesma forma, existe uma carga líquida negativa na solução A a partir da maior concentração de iões cloreto negativos do que os iões sódio positivos. Uma vez que cargas opostas atraem e como cargas repelem, os iões são agora também influenciados por campos eléctricos, bem como por forças de difusão. Por conseguinte, os iões de sódio positivos terão menos probabilidades de viajar para a solução agora mais positiva B e permanecer na solução agora mais negativa A. O ponto em que as forças dos campos eléctricos neutralizam completamente a força devido à difusão é chamado potencial de equilíbrio. Neste ponto, o fluxo líquido do íon específico (neste caso, o sódio) é zero.
Membranas plasmáticasEdit

Cada célula está encerrada numa membrana plasmática, que tem a estrutura de um bocal lipídico com muitos tipos de moléculas grandes nele incorporadas. Por ser feita de moléculas lipídicas, a membrana plasmática tem intrinsecamente uma alta resistividade eléctrica, por outras palavras, uma baixa permeabilidade intrínseca aos iões. Contudo, algumas das moléculas incrustadas na membrana são capazes de transportar activamente iões de um lado da membrana para o outro ou de fornecer canais através dos quais se podem mover.
Em terminologia eléctrica, a membrana plasmática funciona como uma resistência e um condensador combinados. A resistência resulta do facto de a membrana impedir o movimento de cargas através dela. A capacitância surge do facto de o bocal lipídico ser tão fino que uma acumulação de partículas carregadas de um lado dá origem a uma força eléctrica que puxa partículas carregadas em sentido contrário para o outro lado. A capacitância da membrana é relativamente não afectada pelas moléculas que nela estão incorporadas, pelo que tem um valor mais ou menos invariável estimado em cerca de 2 μF/cm2 (a capacitância total de um pedaço de membrana é proporcional à sua área). A condutância de um bocal lipídico puro é tão baixa, por outro lado, que em situações biológicas é sempre dominada pela condutância de vias alternativas fornecidas pelas moléculas incrustadas. Assim, a capacidade da membrana é mais ou menos fixa, mas a resistência é altamente variável.
A espessura de uma membrana de plasma é estimada em cerca de 7-8 nanómetros. Como a membrana é tão fina, não é necessária uma tensão transmembrana muito grande para criar um forte campo eléctrico dentro dela. Os potenciais típicos de membrana em células animais são da ordem dos 100 milivolts (ou seja, um décimo de volts), mas os cálculos mostram que isto gera um campo eléctrico próximo do máximo que a membrana pode suportar – foi calculado que uma diferença de voltagem muito maior do que 200 milivolts poderia causar uma ruptura dieléctrica, ou seja, um arco ao longo da membrana.
Difusão e transporte facilitadosEdit

A resistência de um bico lipídico puro à passagem de iões através dele é muito elevada, mas as estruturas incrustadas na membrana podem melhorar muito o movimento iónico, quer activa quer passivamente, através de mecanismos chamados de transporte facilitado e difusão facilitada. Os dois tipos de estruturas que desempenham os maiores papéis são os canais iónicos e as bombas iónicas, ambos normalmente formados a partir de conjuntos de moléculas de proteínas. Os canais de iões proporcionam passagens através das quais os iões se podem mover. Na maioria dos casos, um canal iónico é permeável apenas a tipos específicos de iões (por exemplo, sódio e potássio, mas não cloreto ou cálcio), e por vezes a permeabilidade varia em função da direcção do movimento iónico. As bombas de iões, também conhecidas como transportadores de iões ou proteínas transportadoras, transportam activamente tipos específicos de iões de um lado da membrana para o outro, por vezes utilizando energia derivada de processos metabólicos para o fazer.
Bombas de iõesEdit
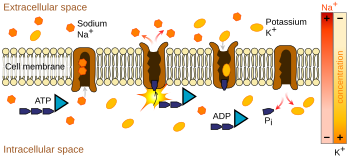
Bombas de iões são proteínas de membrana integral que realizam transporte activo, ou seja, utilizam energia celular (ATP) para “bombear” os iões contra o seu gradiente de concentração. Tais bombas de iões absorvem iões de um lado da membrana (diminuindo a sua concentração ali) e libertam-nos do outro lado (aumentando a sua concentração ali).
A bomba de iões mais relevante para o potencial de acção é a bomba de sódio-potássio, que transporta três iões de sódio para fora da célula e dois iões de potássio para dentro. Como consequência, a concentração de iões de potássio K+ dentro do neurónio é cerca de 20 vezes maior do que a concentração externa, enquanto que a concentração de sódio fora é cerca de nove vezes maior do que a concentração interna. De forma semelhante, outros iões têm concentrações diferentes dentro e fora do neurónio, tais como cálcio, cloreto e magnésio.
Se os números de cada tipo de ião fossem iguais, a bomba de sódio-potássio seria electricamente neutra, mas, devido à troca três por dois, dá um movimento líquido de uma carga positiva de intracelular para extracelular para cada ciclo, contribuindo assim para uma diferença de voltagem positiva. A bomba tem três efeitos: (1) torna a concentração de sódio alta no espaço extracelular e baixa no espaço intracelular; (2) torna a concentração de potássio alta no espaço intracelular e baixa no espaço extracelular; (3) dá ao espaço intracelular uma voltagem negativa em relação ao espaço extracelular.
A bomba de sódio-potássio é relativamente lenta no funcionamento. Se uma célula fosse inicializada com concentrações iguais de sódio e potássio em todo o lado, levaria horas para a bomba estabelecer o equilíbrio. A bomba funciona constantemente, mas torna-se progressivamente menos eficiente à medida que as concentrações de sódio e potássio disponíveis para bombeamento são reduzidas.
As bombas de iões influenciam o potencial de acção apenas estabelecendo a relação relativa das concentrações de iões intracelulares e extracelulares. O potencial de acção envolve principalmente a abertura e fecho de canais de iões e não bombas de iões. Se as bombas de iões forem desligadas pela remoção da sua fonte de energia, ou pela adição de um inibidor como o ouabain, o axon ainda pode disparar centenas de milhares de potenciais de acção antes das suas amplitudes começarem a deteriorar-se significativamente. Em particular, as bombas de iões não desempenham um papel significativo na repolarização da membrana após um potencial de acção.
Outra bomba de iões funcionalmente importante é o permutador de sódio-cálcio. Esta bomba funciona de forma conceptualmente semelhante à bomba de sódio-potássio, excepto que em cada ciclo troca três Na+ do espaço extracelular por um Ca++ do espaço intracelular. Uma vez que o fluxo líquido de carga é para dentro, esta bomba funciona “em descida”, de facto, e portanto não requer qualquer fonte de energia excepto a tensão da membrana. O seu efeito mais importante é bombear o cálcio para fora – também permite um fluxo de sódio para dentro, neutralizando assim a bomba de sódio-potássio, mas, como as concentrações globais de sódio e potássio são muito mais elevadas do que as concentrações de cálcio, este efeito é relativamente insignificante. O resultado líquido do permutador sódio-cálcio é que no estado de repouso, as concentrações intracelulares de cálcio tornam-se muito baixas.
Canais de iõesEditar

Os canais de iões são proteínas de membrana integral com um poro através do qual os iões podem viajar entre o espaço extracelular e o interior da célula. A maioria dos canais são específicos (selectivos) para um íon; por exemplo, a maioria dos canais de potássio são caracterizados por uma razão de selectividade de 1000:1 para potássio sobre sódio, embora os íons potássio e sódio tenham a mesma carga e diferem apenas ligeiramente no seu raio. O poro do canal é tipicamente tão pequeno que os iões têm de passar por ele em ordem de ficheiro único. Os poros dos canais podem ser abertos ou fechados para passagem de iões, embora vários canais demonstrem vários níveis de sub-condutância. Quando um canal está aberto, os iões penetram através do poro do canal, descendo o gradiente de concentração transmembrana para esse ião em particular. A taxa de fluxo iónico através do canal, ou seja, a amplitude da corrente de um canal, é determinada pela condutância máxima do canal e pela força motriz electroquímica para esse ião, que é a diferença entre o valor instantâneo do potencial da membrana e o valor do potencial de inversão.

Um canal pode ter vários estados diferentes (correspondentes a diferentes conformações da proteína), mas cada um desses estados ou é aberto ou fechado. Em geral, os estados fechados correspondem quer a uma contracção do poro – tornando-o intransitável ao ião – quer a uma parte separada da proteína, parando o poro. Por exemplo, o canal de sódio dependente da tensão sofre uma inactivação, na qual uma porção da proteína balança para o poro, selando-o. Esta inactivação desliga a corrente de sódio e desempenha um papel crítico no potencial de acção.
Os canais de íons podem ser classificados pela forma como respondem ao seu ambiente. Por exemplo, os canais iónicos envolvidos no potencial de acção são canais sensíveis à tensão; abrem-se e fecham-se em resposta à tensão através da membrana. Os canais de ligação e os canais de íons formam outra classe importante; estes canais de íons abrem e fecham em resposta à ligação de uma molécula de ligando, como um neurotransmissor. Outros canais de iões abrem e fecham com forças mecânicas. Ainda outros canais iónicos – como os dos neurónios sensoriais – abrem e fecham em resposta a outros estímulos, tais como luz, temperatura ou pressão.
Canais de fugaEditar
Canais de fuga são o tipo mais simples de canal iónico, na medida em que a sua permeabilidade é mais ou menos constante. Os tipos de canais de fuga que têm o maior significado nos neurónios são os canais de potássio e cloreto. Mesmo estes não são perfeitamente constantes nas suas propriedades: Primeiro, a maioria deles são dependentes da tensão, no sentido em que conduzem melhor numa direcção do que na outra (por outras palavras, são rectificadores); segundo, alguns deles são capazes de serem desligados por ligandos químicos, embora não necessitem de ligandos para funcionar.
Canais de ligadura e agregadosEdit
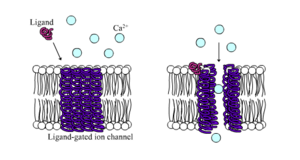
LigandOs canais de iões fechados são canais cuja permeabilidade é grandemente aumentada quando algum tipo de ligante químico se liga à estrutura da proteína. As células animais contêm centenas, se não milhares, de tipos destes. Um grande subconjunto funciona como receptores neurotransmissores – eles ocorrem em locais pós-sinápticos, e o ligante químico que os atravessa é libertado pelo terminal axonal pré-sináptico. Um exemplo deste tipo é o receptor AMPA, um receptor para o glutamato neurotransmissor que quando activado permite a passagem de iões de sódio e potássio. Outro exemplo é o receptor GABAA, um receptor para o neurotransmissor GABA que, quando activado permite a passagem de iões cloreto.
Neurotransmissor receptores são activados por ligandos que aparecem na área extracelular, mas existem outros tipos de canais de liga-agregada que são controlados por interacções do lado intracelular.
Canais dependentes da tensãoEditar
Canais de iões em cadeia, também conhecidos como canais de iões dependentes da tensão, são canais cuja permeabilidade é influenciada pelo potencial da membrana. Formam outro grupo muito grande, com cada membro com uma selectividade iónica particular e uma particular dependência de voltagem. Muitos são também dependentes do tempo – por outras palavras, não respondem imediatamente a uma mudança de voltagem mas apenas após um atraso.
Um dos membros mais importantes deste grupo é um tipo de canal de sódio de tensão que está subjacente ao potencial de acção – estes são por vezes chamados canais de sódio Hodgkin-Huxley porque foram inicialmente caracterizados por Alan Lloyd Hodgkin e Andrew Huxley nos seus estudos vencedores do Prémio Nobel da fisiologia do potencial de acção. O canal é fechado ao nível da tensão de repouso, mas abre abruptamente quando a tensão excede um certo limiar, permitindo um grande afluxo de iões de sódio que produz uma mudança muito rápida no potencial de membrana. A recuperação de um potencial de acção depende, em parte, de um tipo de canal de potássio de tensão, fechado ao nível da tensão de repouso mas aberto em consequência da grande mudança de tensão produzida durante o potencial de acção.
Potencial de reversãoEditar
O potencial de reversão (ou potencial de equilíbrio) de um ião é o valor da tensão transmembrana em que as forças difusoras e eléctricas contrabalançam, de modo a que não haja fluxo líquido de iões através da membrana. Isto significa que a tensão transmembrana se opõe exactamente à força de difusão do íon, de modo a que a corrente líquida do íon através da membrana seja zero e imutável. O potencial de reversão é importante porque dá a tensão que actua nos canais permeáveis a esse ião – por outras palavras, dá a tensão que o gradiente de concentração do ião gera quando actua como uma bateria.
O potencial de equilíbrio de um determinado ião é normalmente designado pela notação Eion.O potencial de equilíbrio para qualquer ião pode ser calculado usando a equação de Nernst. Por exemplo, o potencial de reversão dos iões de potássio será o seguinte:
E e q , K + = R T z F ln o i , {\i1}displaystyle E_{eq,K^{+}}={\i}{\i1}frac {RT}{zF}}ln {\i}{\i}{\i},}

where
- Eeq,K+ é o potencial de equilíbrio do potássio, medido em volts
- R é a constante universal do gás, igual a 8,314 joules-K-1-mol-1
- T é a temperatura absoluta, medida em kelvins (= K = graus Celsius + 273.15)
- z é o número de cargas elementares do ião em questão envolvidas na reacção
- F é a constante de Faraday, igual a 96,485 coulombs-mol-1 ou J-V-1-mol-1
- o é a concentração extracelular de potássio, medida em mol-m-3 ou mmol-l-1
- i é a concentração intracelular de potássio
p>P>Even se dois iões diferentes tiverem a mesma carga (i.e., K+ e Na+), ainda podem ter potenciais de equilíbrio muito diferentes, desde que as suas concentrações exteriores e/ou interiores sejam diferentes. Tomemos, por exemplo, os potenciais de equilíbrio de potássio e sódio nos neurónios. O potencial de equilíbrio de potássio EK é de -84 mV com 5 mM de potássio no exterior e 140 mM no interior. Por outro lado, o potencial de equilíbrio de sódio, ENa, é aproximadamente +66 mV com aproximadamente 12 mM de sódio no interior e 140 mM no exterior.
Alterações do potencial de membrana durante o desenvolvimentoEdit
Um potencial de membrana de repouso do neurónio muda realmente durante o desenvolvimento de um organismo. Para que um neurónio possa eventualmente adoptar a sua função adulta completa, o seu potencial deve ser rigorosamente regulado durante o desenvolvimento. À medida que um organismo progride através do desenvolvimento, o potencial de membrana de repouso torna-se mais negativo. As células giais estão também a diferenciar-se e a proliferar à medida que o desenvolvimento progride no cérebro. A adição destas células gliais aumenta a capacidade do organismo de regular o potássio extracelular. A queda no potássio extracelular pode levar a uma diminuição do potencial de membrana de 35 mV.
Excitabilidade celularEditar
Excitabilidade celular é a alteração do potencial de membrana necessária para respostas celulares em vários tecidos. A excitabilidade celular é uma propriedade que é induzida durante a embriogénese precoce. A excitabilidade de uma célula foi também definida como a facilidade com que uma resposta pode ser desencadeada. Os potenciais de repouso e limiar formam a base da excitabilidade celular e estes processos são fundamentais para a geração de potenciais graduados e de acção.
Os reguladores mais importantes da excitabilidade celular são as concentrações extracelulares de electrólitos (i.e. Na+, K+, Ca2+, Cl-, Mg2+) e proteínas associadas. As proteínas importantes que regulam a excitabilidade celular são os canais de iões de tensão, transportadores de iões (por exemplo, Na+/K+-ATPase, transportadores de magnésio, transportadores de base ácida), receptores de membrana e canais cíclico-nucleótidos activados por hiperpolarização. Por exemplo, os canais de potássio e os receptores com sensibilidade ao cálcio são importantes reguladores da excitabilidade em neurónios, miócitos cardíacos e muitas outras células excitáveis como as astrocitos. O ião de cálcio é também o segundo mensageiro mais importante na sinalização das células excitáveis. A activação de receptores sinápticos inicia mudanças duradouras na excitabilidade neuronal. A tiróide, adrenal e outras hormonas também regulam a excitabilidade celular.
Muitos tipos de células são considerados como tendo uma membrana excitável. Células excitáveis são neurónios, miócitos (cardíacos, esqueléticos, lisos), células endoteliais vasculares, células justaglomerulares, células intersticiais de Cajal, muitos tipos de células epiteliais (por exemplo células beta, células alfa, células delta, células enteroendócrinas), células gliais (por exemplo astrocitos), células mecanorreceptoras (por exemplo células capilares e células Merkel), células quimiorreceptoras (por exemplo células glomosas, receptores gustativos), algumas células vegetais e possivelmente células imunitárias. Os astrócitos apresentam uma forma de excitabilidade não eléctrica baseada em variações intracelulares de cálcio relacionadas com a expressão de vários receptores através dos quais podem detectar o sinal sináptico. Nos neurónios, existem diferentes propriedades de membrana em algumas partes da célula, por exemplo, a excitabilidade dendrítica confere aos neurónios a capacidade de detecção por coincidência de entradas separadas espacialmente.
Circuito equivalenteEdit
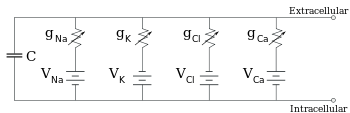
Electrofisiologistas modelam os efeitos das diferenças de concentração iónica, canais iónicos e capacitância de membrana em termos de um circuito equivalente, que se destina a representar as propriedades eléctricas de uma pequena mancha de membrana. O circuito equivalente consiste num condensador em paralelo com quatro vias, cada uma delas constituída por uma bateria em série com uma condutância variável. A capacitância é determinada pelas propriedades do bocal lipídico, e é tomada para ser fixa. Cada uma das quatro vias paralelas provém de um dos iões principais, sódio, potássio, cloreto, e cálcio. A tensão de cada via iónica é determinada pelas concentrações do ião em cada lado da membrana; ver a secção sobre o potencial de reversão acima. A condutância de cada via iónica em qualquer momento é determinada pelos estados de todos os canais iónicos que são potencialmente permeáveis a esse ião, incluindo os canais de fuga, os canais de ligação e os canais iónicos de ligação e os canais iónicos de tensão.
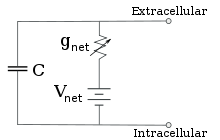
Para concentrações fixas de iões e valores fixos de condutância de canais iónicos, o circuito equivalente pode ser ainda mais reduzido, utilizando a equação de Goldman, como descrito abaixo, a um circuito contendo uma capacitância em paralelo com uma bateria e uma condutância. Em termos eléctricos, este é um tipo de circuito RC (circuito de capacidade de resistência), e as suas propriedades eléctricas são muito simples. Partindo de qualquer estado inicial, a corrente que flui através da condutância ou da capacitância decai com um curso temporal exponencial, com uma constante de tempo de τ = RC, onde C é a capacitância do adesivo de membrana, e R = 1/gnet é a resistência da rede. Para situações realistas, a constante de tempo encontra-se normalmente na gama de 1-100 milissegundos. Na maioria dos casos, as alterações na condutância dos canais iónicos ocorrem numa escala de tempo mais rápida, pelo que um circuito RC não é uma boa aproximação; no entanto, a equação diferencial utilizada para modelar um patch de membrana é geralmente uma versão modificada da equação do circuito RC.