Zellbiologie 07: Mikrotubuli und Zellteilung
Dies sind Notizen zur Vorlesung 7 des Kurses Zellbiologie von Harvard Extension.
In der Vorlesung 6 wurden Mikrotubuli eingeführt, und in dieser Vorlesung wird ihre Rolle bei der Zellteilung besprochen. Hier ist ein Einführungsvideo:
Überblick über den Zellzyklus
Der Zellzyklus – der Prozess der Zellteilung und Replikation – wird durch eine Reihe von biochemischen Schaltern gesteuert, die man das Zellzyklus-Kontrollsystem nennt.
Der Zellzyklus ist in Phasen unterteilt, die wiederum in Phasen unterteilt sind – die Leute werden sich auf die „4 Phasen“ beziehen, aber eigentlich sind es 5, und die Leute benutzen auch andere Wörter, um diese Phasen zusammenzufassen, und andere Wörter, um sie zu unterteilen. Ich habe mein Bestes getan, um die Beziehung zwischen diesen Begriffen in der folgenden Tabelle zusammenzufassen. (modifiziert/erweitert aus Wikipedia):
| Allgemeinste Gruppierung | die vermeintlichen „4 Phasen“ | Unterphasen | nicht-.teilend | Lücke 0 (G0) |
|---|---|---|
| Interphase | Lücke 1 (G1) | G1a R G1b |
| Synthese (S) | ||
| Gap 2 (G2) | ||
| Mitose | Mitose (M) | Prophase Prometaphase Metaphase Anaphase Telophase Zytokinese |
Der Inhalt der einzelnen Phasen ist in diesem hervorragenden Wikimedia-Commons-Bild von Kelvinsong schön zusammengefasst:

Die sich am schnellsten teilenden menschlichen Zellen können einen Zellzyklus in etwa 24 Stunden abschließen (G1: 9h, S: 10h, G2: 4h, M: 30 min). Hefe kann einen Zyklus in 30 Minuten abschließen, und die sich am schnellsten teilenden Drosophila-Zellen brauchen nur 8 Minuten.
Zu den Hauptkontrolleuren dieses Prozesses gehören die Cycline, die die Cyclin-abhängige Kinase oder CDK regulieren. Zur Erinnerung: Kinasen sind Proteine, die andere Proteine phosphorylieren. Die Phosphorylierung ihrer Ziele durch die CDK ermöglicht den Ablauf der Mitose. Um genau zu sein, ist der reifungsfördernde Faktor oder MPF ein obligater heterodimerer Komplex, der aus Cyclin B und CDK besteht und nur dann seine phosphorylierende Wirkung entfaltet, wenn beide Proteine vorhanden sind.
Rolle der Mikrotubuli
Mikrotubuli sind während des gesamten Zellzyklus von entscheidender Bedeutung – sie organisieren zelluläre Komponenten und teilen sie in zwei Teile. Hier sind eine Reihe von Videos des Zellzyklus, die die Rolle der Mikrotubuli hervorheben:
Bei Tieren haben ruhende Zellen und sogar Zellen in der Interphase normalerweise nur ein MTOC, genannt Zentrosom, das als zentraler Knotenpunkt für alle Mikrotubuli in der Zelle dient. Ein Zentrosom besteht aus zwei Zentriolen, wie unten gezeigt (wieder mit Dank an Kelvinsong):

Die beiden Zentriolen lösen sich voneinander und replizieren sich während der S-Phase und trennen sich dann, um während der M-Phase entgegengesetzte „Pole“ der Zelle zu bilden, so dass es jetzt zwei MTOCs gibt, von denen jedes schließlich das einzige MTOC einer neuen Zelle sein wird (ein weiteres Bild von Chef Kelvinsong):
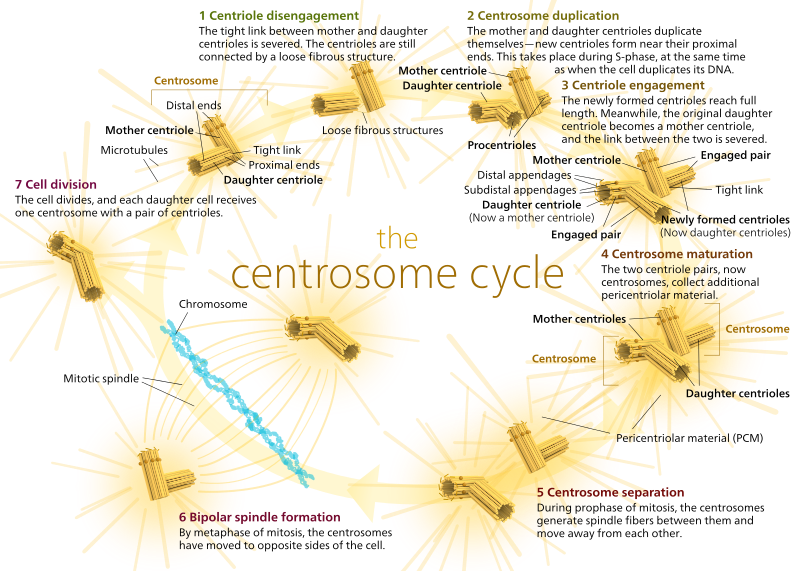
Während der Mitose haben Sie also die beiden „Pole“ der Zelle, jeweils mit Mikrotubuli, die am (-) Ende verankert sind und deren (+) Enden sich überlappen und in die Mitte der Zelle zeigen, wie hier gezeigt (Wikimedia Commons Bild von Lordjuppiter):
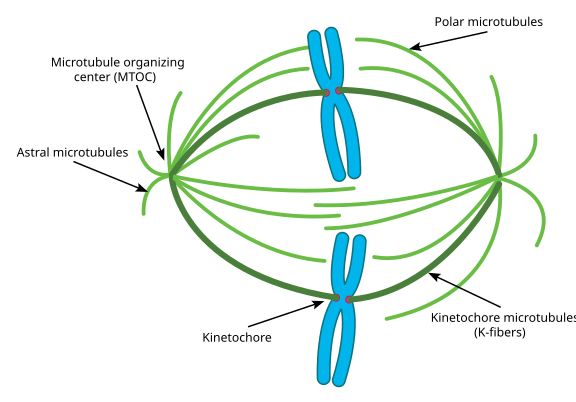
Das Ganze nennt man Spindelapparat, und den Bereich, in dem sich die Mikrotubuli der beiden MTOCs überlappen, nennt man die „Zone der Interdigitation“. Manchmal hört man, dass jedes MTOC und seine igelartige Anordnung von Mikrotubuli als „mitotische Aster“ bezeichnet wird.
Mikrotubuli in diesem Stadium werden in drei Kategorien eingeteilt:
- Astrale Mikrotubuli zeigen nach außen, in Richtung Zellrinde, um den gesamten Spindelapparat entlang der Achse der Zellteilung zu verankern.
- Kinetochore Mikrotubuli befestigen sich am Kinetochor der Chromatiden.
- Polare Mikrotubuli, die parallel zueinander, aber in entgegengesetzte Richtungen ausgerichtet sind, sind entscheidend für das Auseinanderschieben des Spindelapparats während der Mitose. (Tatsächlich sind polare Mikrotubuli auch schon früher vorhanden und helfen, die Zentrosomen während der Prophase auseinander zu schieben).
Wenn Sie Fotos gegenüber Diagrammen bevorzugen, sehen Sie hier, wie der gesamte Spindelapparat aussieht, mit Chromatiden in blau, Mikrotubuli in grün und den Kinetochoren als rote Punkte:

Mikrotubuli werden während der Mitose viel dynamischer: Mehr Gamma-Tubulin fördert die leichtere Nukleation, aber XMAP215, ein Mikrotubuli-Stabilisator, wird während der Mitose phosphoryliert und damit inaktiviert, so dass Kinesin-13 frei wird, um die Mikrotubuli zu katastrophieren. Ein Vermögen wird schnell gemacht und verloren. Die Halbwertszeit eines Mikrotubulus während der Mitose beträgt etwa 15 Minuten, im Vergleich zu 30 Minuten während der Interphase. Man studiert die Mikrotubuli-Dynamik mit Hilfe von FRAP: Man fügt ein fluoreszierendes Mikrotubuli hinzu, bleicht es und schaut, wie schnell der Wiederzusammenbau stattfindet, basierend darauf, wie schnell die Fluoreszenz wieder erscheint.
Kinesin-5 hat zwei polare Köpfe, die sich an gegenüberliegende Mikrotubuli binden und versuchen, zum (+)-Ende der Mikrotubuli zu wandern. Dies schiebt die beiden Mikrotubuli auseinander und liefert die treibende Kraft für die Trennung der MTOCs.
Zentromerische DNA hat eine niedrige Informationsentropie und spezielle Histone, die sich von anderem Chromatin unterscheiden. Zentromere sind ein Teil des Genoms, der bei der Next-Gen-Sequenzierung fast nie erfasst wird, selbst bei sehr großer Tiefe. Das liegt daran, dass die Zentromere einem anderen Zweck dienen als ein Großteil des restlichen Genoms: Die Sequenz dort ist günstig für die Interaktion mit zentromerischen Proteinen und die Kinetochor-Anbindung. Cohesine sind Proteine, die die beiden Schwesterchromatiden zusammenhalten. Bei den Kinetochor-Proteinen sprechen wir von zwei Schichten, dem inneren Kinetochor und dem äußeren Kinetochor.
Während der Prometaphase bewegen sich die Chromosomen hin und her. Kinesine verankern die Chromosomen an den Kinetochor-Mikrotubuli jenseits der Spitze, wo Kinesin-13 die Mikrotubuli depolymerisiert, unterstützt durch einen Mangel an verfügbaren Tubulin-Dimeren. Eine Kombination aus Motorproteinen, Mikrotubuli-interagierenden Proteinen und Treadmilling dient der Bewegung der Chromosomen. Währenddessen arbeiten Dynein und Dynactin – Motorproteine, die zum (-)Ende hin laufen – an den astralen Mikrotubuli und ziehen die MTOCs in Richtung Zellperipherie. In der Metaphase richten sich die Chromatiden entlang der „Metaphasenplatte“ aus.
Während dieses Prozesses löst sich die Kernhülle auf, so dass der Kernimport irrelevant wird. Ran-GEF lokalisiert sich in der Nähe der Chromosomen und erzeugt hohe Konzentrationen von Ran-GTP, das Energie für einige notwendige Prozesse bereitstellt (?).
Zellen haben irgendeinen Mechanismus, um die Spannung in den Mikrotubuli zu erkennen, die ihre Anheftungschromatiden anzeigt, bevor die Mitose fortgesetzt werden kann. Sicherzustellen, dass jedes Chromatid richtig verankert ist, ist entscheidend für die Vermeidung von Aneuploidie.
Neben den Mikrotubuli spielen übrigens auch andere Zytoskelettelemente eine wichtige Rolle im Zellzyklus. Bei der Zytokinese bildet Aktin einen kontraktilen Ring und spannt mit Hilfe von Myosin-II-Motorproteinen die Zelle in zwei Hälften.
Bedeutung von Modellorganismen
Die Entdeckung von zellzyklusregulatorischen Prozessen stützte sich stark auf einige nette Eigenschaften bekannter Modellorganismen.
Saccharomyces cerevisiae (Knospende Hefe) und Schizosaccharomyces pombe (Spalthefe) können als Haploide oder Diploide existieren. Das ist wichtig, weil in der haploiden Phase eine Mutation ein Gen ausschalten kann – man muss nicht beide Allele treffen. Und in Hefe sind viele Mutationen, besonders in den Cdc__-Genen (Zellteilungssteuerung), temperaturabhängig, wobei ein Protein mit einer Missense-Mutation bei „permissiven“ Temperaturen noch richtig funktionieren kann, aber bei „nicht-permissiven“ Temperaturen seine native Funktion verliert. Dadurch ist es möglich, den Knockout-Phänotyp (bei der nicht-permissiven Temperatur) zu studieren, während die Organismen (bei der permissiven Temperatur) einfach vermehrt werden können. Das gesamte S. cerevisiae-Genom ist in Form von Plasmidbibliotheken verfügbar, wodurch es möglich ist, zu screenen, welches Plasmid den Phänotyp einer bestimmten Mutante rettet. Auf diese Weise wurden viele der Gene entdeckt, die den Zellzyklus regulieren.
In S. cerivisiae ist die Knospung Teil der Phase G1, und sobald die Tochterzelle eine bestimmte Größe erreicht, in einem Moment, der „START“ genannt wird, sind die beiden verpflichtet, in S einzutreten und den Zellzyklus schließlich zu beenden. Säugetierzellen haben ihren eigenen Commitment-Punkt, den Restriktionspunkt oder R, in G1, der analog zu START ist.
Temperaturempfindliche Cdc28-Mutanten knospen nicht bei der nicht-permissiven Temperatur. Das Cdc28-Gen kodiert das Hefe-Homolog unserer Cyclin-abhängigen Kinase (CDK), die, wenn und nur wenn sie mit Cyclin komplexiert ist, andere Proteine phosphorylieren kann, um deren Beteiligung an Zellzyklusphasen zu regulieren. Temperatursensitive Mutanten bleiben bei der nicht-permissiven Temperatur stecken und können nicht in die S-Phase übergehen. Stattdessen verhalten sie sich wie Wildtyp-Zellen, denen die Nährstoffe entzogen werden: Sie wachsen groß genug, um die START-Phase zu durchlaufen, setzen dann aber nicht fort.
Xenopus (eine Froschart) erwies sich als entscheidend für das Verständnis des Zellzyklus, da seine Fortpflanzung eine sehr große Anzahl von Zellen umfasst (d. h. genügend Ausgangsmaterial für Western Blots usw.), die perfekt synchronisiert sind (d. h. alle befinden sich zum gleichen Zeitpunkt in der gleichen Phase des Zellzyklus. (Im Vergleich zu Hefe zum Beispiel, wo nicht alle Zellen zur gleichen Zeit in der gleichen Phase sind). Auch das Ei selbst ist groß und leicht zu bearbeiten, und der Befruchtung folgen mehrere Zellzyklen. Bei Fröschen beginnen die Eier mit der meiotischen Teilung, verharren dann aber 8 Monate lang in der G2-Phase, während sie an Größe zunehmen und Dinge ansammeln, die bei der Befruchtung für das Wachstum benötigt werden.
Zwischenfilamente
Zusätzlich zu den Mikrofilamenten und Mikrotubuli haben eukaryotische Zellen auch eine Vielzahl von „anderen“ Zytoskelettproteinen, die Zwischenfilamente (IFs) genannt werden. Obwohl sie vielfältiger sind als Mikrofilamente und Mikrotubuli, sind IFs nicht nur ein Sammelbegriff für „jedes andere Filament“ – vielmehr sind sie eine Gruppe verwandter Proteine. Sie erstrecken sich im Allgemeinen durch das Zytoplasma und die innere Kernhülle, sind unpolar und haben keine mit ihnen assoziierten Motorproteine. Sie haben eine große Zugfestigkeit und sind sehr stabil, mit einer langsamen Austauschrate und wenig Abbau, obwohl Phosphorylierung ihre Demontage fördern kann. Hier sind einige populäre Beispiele:
- Keratine finden sich in Epithelzellen, Mesodermzellen und Neuronen. Sie sorgen für Festigkeit und kommen in sauren und basischen Formen vor. Jedes kann einen eigenen Strang bilden, aber die meisten IFs bestehen aus zwei Strängen – einem basischen und einem sauren, die gewissermaßen umeinander gedreht sind. Haare und Nägel bestehen aus „hartem“ Keratin, das reich an Cystein für Disulfidbindungen ist, die für die immense Stärke sorgen. Dauerwellen und Glättungen beruhen auf der Reduzierung der Disulfidbindungen, der Umformung des Haares und der anschließenden Neuformung der Disulfidbindungen. Sie haben auch „weiches“ Keratin in Ihrer Haut.
- Desmine wie Vimentin finden sich in mesenchymalen Zellen (Knochen, Knorpel und Fett).
- Neurofilamente befinden sich in neuronalen Axonen und regulieren deren Durchmesser, was wiederum die Geschwindigkeit der Ausbreitung von Aktionspotentialen bestimmt.
- Lamine sind sowohl die am weitesten verbreiteten als auch die vermutlich ähnlichsten phylogenetischen Vorfahren aller anderen IFs. Sie bieten strukturelle Unterstützung für die Kernmembran. Sie könnten dabei helfen, die Kernporenkomplexe zu verteilen und auch die DNA zu organisieren.
Zuletzt noch ein zusammenfassendes Video: